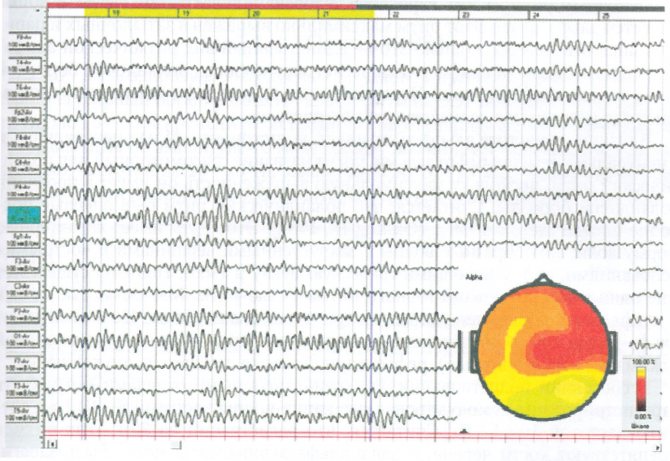
Рис. 1. Альфа-паттерн ЭЭГ модулированный в «веретена» ритм частотой 10 Гц, максимально выраженный в затылочных отведениях и демонстрирующий выраженный фронтоокципитальный градиент амплитуды. В правой части: топограмма распределения альфа-ритма по мощности, эпоха анализа отмечена двумя маркерами
При соблюдении стандартных условий регистрации (закрытые глаза, состояние спокойного бодрствования, эмоциональный комфорт, отсутствие внешних раздражителей) у большинства (85-90%) взрослых здоровых людей в норме на ЭЭГ регистрируется альфа-паттерн, представляющий собой организованный альфа-ритм, четко модулированный в веретена амплитудой от 40 до 60— 80 мкВ (рис. 1).
Форма волн правильная синусоидальная: не «заострена», не искажена асимметрией восходящей и нисходящей фаз. На нормальной ЭЭГ альфа-ритм занимает до 80—90 % времени регистрации (индекс ритма), изредка прерываясь эпизодами быстроволновой активности, очевидно, в связи с собственной психической (мыслительной) деятельностью обследуемого.
Альфа-ритм максимально представлен в париетальных и затылочных отделах обоих полушарий. Выражена зональность распределения альфа-ритма: его амплитуда и частота постепенно убывают от затылка к центральным и лобным отделам мозга. В лобных отведениях регистрируется слабовыраженный низкоамплитудный немодулированный альфа-ритм, занимающий не более 50 % времени регистрации, чередуясь с периодами регистрации низкочастотной бета-активности, сравнимой с альфа-ритмом по амплитуде. В височных отведениях регистрируется не модулированный в веретена альфа-ритм, как правило, с меньшей амплитудой, чем в центральных отведениях. Медленные волны во всех отведениях практически не представлены.
Симметричность, пусть и относительная, настолько характерна для нормальной ЭЭГ, что является одним из критериев оценки функционального состояния мозга. В норме асимметрия по амплитуде не превышает 30 %. Частотная межполушарная асимметрия не должна быть более 1 Гц. Естественно, при этом должна быть полная уверенность в идентичности установки электродов над обоими полушариями. Следствием функциональной асимметрии мозга, связанной у большинства людей с преобладанием активности левого (доминантного) полушария, является некоторое преобладание по амплитуде альфа-ритма в правом (!) полушарии. Считается, что левое полушарие более тесно связано с активирующими мезэнцефальными структурами мозга, а правое — с синхронизирующими диэнцефальными структурами.
Механизмы формирования нормальной ЭЭГ
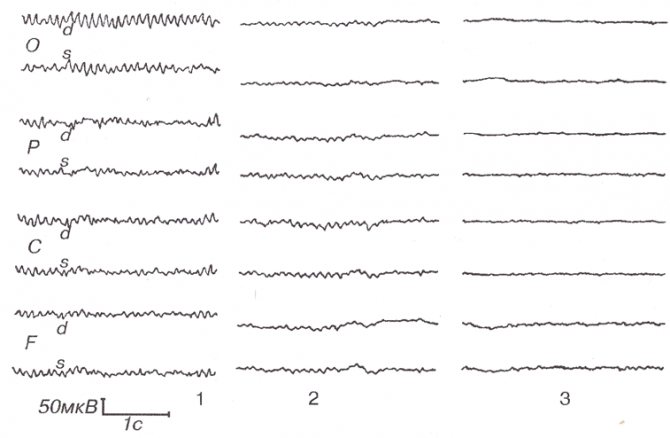
Рис. 2. Варианты нормальной ЭЭГ
1 — наиболее часто встречающийся тип ЭЭГ, 2 — умеренно сниженная амплитуда ЭЭГ; 3 — низкоамплитудная ЭЭГ.
Нормальная ЭЭГ бодрствования зависит от механизмов, определяющих уровень функциональной активности всего мозга. Особенностями этих систем являются их срединное расположение в мозге, синхронизация активности в двух полушариях на всех подкорковых уровнях, а также диффузная и симметричная связь этих систем с корой (см. рис. 2). Вследствие этого ЭЭГ в существенной степени однородна для всего мозга и симметрична. Следует, однако, отметить, что, несмотря на диффузный характер влияний срединных структур мозга на вышележащие уровни, функциональная и морфологическая неоднородность коры приводит к определенным и существенным особенностям электрической активности различных областей мозга. Тем не менее, вследствие достаточно постепенного перехода одних функциональных зон коры в другие, смена типов ЭЭГ, присущих отдельным областям мозга, по протяженности коры происходит постепенно.
Симметричность настолько характерна для нормальной ЭЭГ, что является одним из существенных критериев диагностики. Практически вариантом нормы можно считать ЭЭГ, на которой значение асимметрии составляет не более 50% амплитуды сравниваемых записей. Естественно, при этом должна быть полная уверенность в идентичности установки и коммутации электродов в обоих полушариях.
Виды активности человеческого мозга, фиксируемые при записи ЭЭГ
Основными видами активности, которые записываются в ходе процедуры и впоследствии подвергают интерпретации, а также дальнейшему изучению считаются волновые частота, амплитуда и фаза.
Частота
Показатель оценивается количеством волновых колебаний за секунду, фиксируется цифрами, и выражается в единице измерения – герцах (Гц). В описании указывается средняя частота изучаемой активности. Как правило, берется 4-5 участков записи длительностью1с, и рассчитывается число волн на каждом временном отрезке.
Амплитуда
Данный показатель – размах волновых колебаний эклектического потенциала. Измеряется расстоянием между пиками волн в противоположных фазах и выражается в микровольтах (мкВ). Для замера амплитуды применяется калибровочный сигнал. Если, к примеру, калибровочный сигнал при напряжении 50 мкВ определяется на записи высотой 10 мм, то 1 мм будет соответствовать 5 мкВ. В расшифровке результатов дается интерпретациям наиболее частым значениям, полностью исключая редко встречающиеся.
Фаза
Значение этого показателя оценивает текущее состояние процесса, и определяет его векторные изменения. На электроэнцефалограмме некоторые феномены оцениваются количеством содержащихся в них фаз. Колебания подразделяются на монофазные, двухфазные и полифазные (содержащие более двух фаз).
Ритмы мозговой деятельности
Понятием «ритм» на электроэнцефалограмме считается тип электрической активности, относящийся к определенному состоянию мозга, координируемый соответствующими механизмами. При расшифровке показателей ритма ЭЭГ головного мозга вносятся его частота, соответствующая состоянию участка мозга, амплитуда, и характерные его изменения при функциональных сменах активности.
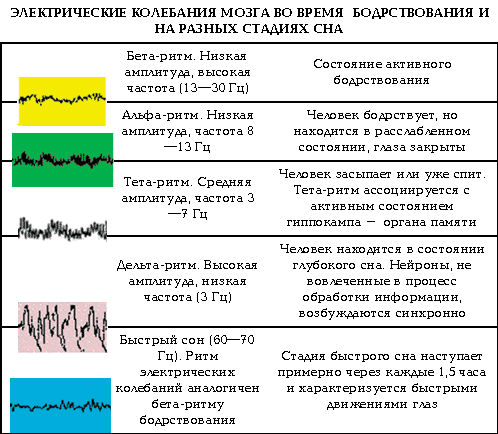
Характеристики ритмов головного мозга зависят от того, в бодрствовании или в состоянии сна находится обследуемый
Роль альфа-ритма в нормальной ЭЭГ
Альфа-ритм регистрируется на ЭЭГ в состоянии спокойного бодрствования, при закрытых глазах и отсутствии внешних сенсорных воздействий. Любая неспецифическая активация: открывание глаз, вспышка света, тональный щелчок, активная мыслительная деятельность, — вызывают блокаду альфа-ритма и реакцию десинхронизации на ЭЭГ. В норме блокада альфа-ритма должна быть полной, т. е. должна происходить полная редукция альфа-активности и «замена» ее на быстроволновые низкоамплитудные осцилляции, и одномоментной во всех отведениях, в которых регистрировался альфа-ритм. При повторном закрывании глаз исходный альфа-паттерн должен восстанавливаться в течение 0,3—1,0 с. Высокая реактивность является одним из существенных признаков нормального альфа-ритма.
Описанный альфа-паттерн представляет собой так называемую «идеальную норму» на ЭЭГ. Такой хрестоматийный вариант регистрируется не всегда. Варианты альфа-паттерна могут характеризоваться сглаженным зональным распределением альфа-ритма, когда градиент амплитуды не столь выражен, и альфа-активность остается достаточно выраженной в передних отделах мозга. Возможно снижение модуляции альфа-веретен. При снижении индекса альфа-ритма могут регистрироваться либо медленноволновая активность, либо активность бета-1 диапазона. При этом индекс альфа-ритма остается достаточно высоким (больше 60—70 %), а амплитуда других компонентов не превышает половины амплитуды альфа-волн. Все перечисленные изменения ЭЭГ трактуются как варианты нормы.
Таким образом, у подавляющего большинства взрослых здоровых обследуемых в стандартных условиях при закрытых глазах на ЭЭГ регистрируется организованный альфа-паттерн, отражающий сбалансированное состояние синхронизирующих и десинхронизирующих систем генерации биоэлектрической активности. Представленность этого типа активности в популяции по результатам различных исследований оценивается на уровне от 60 до 80—90 %.
«Плоская» ЭЭГ
Рис. 3. «Плоская» ЭЭГ
У 10—20 % здоровых обследуемых не удается зарегистрировать альфа-паттерн. Регулярный организованный альфа-ритм на ЭЭГ практически отсутствует. Около 10—20 % времени регистрируется нерегулярная альфа-активность, не превышающая по амплитуде 10 мкВ. На ЭЭГ доминирует бета-активность, как правило, частотой до 20—25 Гц, амплитудой до 10—15 мкВ. Характерный вид такой активности позволил обозначить паттерн как «плоская» ЭЭГ, а тип ЭЭГ называют низкоамплитудными (рис. 3).
«Плоская» ЭЭГ является вариантом нормального состояния биоэлектрической активности. Тем не менее дифференцировать такой паттерн от патологических типов, например от диффузной тахиаритмии при выраженном атеросклеротическом поражении сосудов головного мозга, конечно же, непростая задача. «Плоская» ЭЭГ оценивается как вариант нормы при условии исключения патологических процессов в ЦНС.
Подробно об энцефалограмме
Суть обследования заключается в фиксации электрической активности нейронов структурных образований головного мозга. Электроэнцефалограмма – это своеобразная запись нейронной деятельности на специальной ленте при использовании электродов. Последние закрепляются на участки головы и регистрируют активность определенного участка мозга.
Активность человеческого мозга напрямую определяется работой его срединных образований – переднего мозга и ретикулярной формации (связующего нейронного комплекса), обуславливающих динамику, ритмичность и построение ЭЭГ. Связующая функция формации определяет симметричность и относительную идентичность сигналов между всеми структурами мозга.
Строение головного мозга, на основании этих данных специалист расшифровывает диагностику
Процедура назначается при подозрениях на различные нарушения структуры и деятельности ЦНС (центральной нервной системы) – нейроинфекции, такие как менингит, энцефалит, полиомиелит. При данных патологиях изменяется активность мозговой деятельности, и это сразу же можно диагностировать на ЭЭГ, а в дополнение установить локализацию пораженного участка. ЭЭГ проводится на основании стандартного протокола, в котором фиксируются снятие показателей при бодрствовании или сне (у младенцев), а также с применением специализированных тестов.
К основным тестам относятся:
- фотостимуляция – воздействие на закрытые глаза яркими вспышками света;
- гипервентиляция – глубокое редкое дыхание на протяжении 3-5 минут;
- открытие и закрытие глаз.
Эти тесты считаются стандартными и их применяют при энцефалограмме головного мозга и взрослым и детям любого возраста, и при различных патологиях. Существует еще несколько дополнительных тестов, назначающихся в отдельных случаях, таких как: сжатие пальцев в так называемый кулак, нахождение 40 минут в темноте, лишение сна на определенный период, мониторинг ночного сна, прохождение психологических тестов.
Данные тесты определяются неврологом и добавляются к основным, проводимым в ходе обследования, когда врачу необходимо оценить конкретные функции мозга.
Низкоамплитудные ЭЭГ
Низкоамплитудные ЭЭГ, по современным данным, указывают на преобладание в мозге десинхронизирующих влияний. Этот тип ЭЭГ связан с аутосомно-доминантным геном и формируется постепенно в процессе созревания. До 20 лет он наблюдается чрезвычайно редко1. Представление о том, что этот вариант ЭЭГ связан с десинхронизацией и, соответственно, с преобладанием активирующих восходящих неспецифических систем, согласуется с некоторыми данными психологических исследований. Показано, что низкоамплитудные ЭЭГ коррелируют с повышенной поведенческой активностью, тенденцией к независимости, с агрессивностью, повышенной психической возбудимостью, в то время как высокоамплитудные ЭЭГ характерны для лиц пассивного, зависимого, рецептивного, спокойного типа23. Таким образом, низкоамплитудные ЭЭГ являются вариантом нормы. Следует, однако, отметить, что при некоторых типах нарушений в области нижнего и среднего ствола мозга также возникает уплощение ЭЭГ, внося определенные трудности в клиническую оценку этих кривых.
Особенности проведения
Данный метод исследования считается высокоэффективной техникой для уточнения патологических процессов в мозге и позволяет точно поставить диагноз.
Цель диагностики:
- Оценка периода бодрствования и сна;
- Динамика проводимой терапии;
- Локализация воспалительного очага;
- Выявление патологических изменений.
Фиксирование результатов ЭЭГ ребенка возможно как в состоянии покоя, так и с различными пробами:
- Запись результатов в состоянии сна (фоновая кривая импульсов нейронов мозга);
- Активизация мозговых структур при переходе из состояния покоя в режим деятельности (диагност в установленные промежутки времени говорит, когда необходимо открыть и закрыть глаза);
- Применение пробы — гипервентиляции (пациент по команде диагноста глубоко вдыхает и выдыхает). Данная проба способна выявить новообразования и эпилептические скрытые признаки;
- Диагностика с использованием фотостимуляции (перед ребенком включается лампа с заданным ритмом на включение и выключение, глаза во время пробы закрыты). Данная проба оценивает реакцию на раздражитель при нарушениях психического и речевого развития. Применяется так же при эпилепсии и судорожном синдроме.
ЭЭГ мозга у детей, возможно, проводить, диагностируя импульсы в ночное время, или методом депревации ночного сна (ребенку длительное время не дают спать, с дальнейшей записью активности мозга во время покоя), но данная проба назначается крайне редко.
От редакции : Симптомы, причины и методика лечения невралгии
Алгоритм выполнения ЭЭГ маленьким пациентам в основном определяется их возрастом. Так, малыши до года во время прохождения исследования лежат у мамы на руках либо их укладывают на пеленальный столик
Очень важно, чтобы ребенок на протяжении диагностического мониторинга спал. В случае если малыш капризничает, его сначала необходимо успокоить и подождать, пока он заснет
Сеанс ЭЭГ, как правило, длится не более 20 минут. Цель исследования – зарегистрировать БЭА мозга ребенка в спокойном состоянии. Деткам постарше могут добавляться более сложные диагностические техники, такие как функциональные пробы различного характера.
Необходимо, чтобы обследуемый во время диагностирования разместился на кушетке в положении полулежа. Голова не должна наклоняться вперед, иначе полученные материалы могут быть недостоверны. Персонал диагностического кабинета и родители должны внимательно следить за состоянием маленького пациента, чтобы ему было комфортно и он смог полностью расслабиться.
Стандартная процедура энцефалографии для ребенка старше одного года, способного выполнять указания врача, состоит из следующих этапов. Первой осуществляется запись «фоновой кривой», что показывает БЭА в состоянии покоя. Затем проводятся функциональные пробы, позволяющие отследить патологические изменения при смене рода деятельности.
Изменения ЭЭГ в цикле «бодрствование — сон»
➥ Основная статья: Сон: ЭЭГ диагностика нарушений сна
Поскольку ЭЭГ связана с системами, определяющими уровень функциональной активности мозга, можно установить определенное соответствие между характером ЭЭГ и функциональным состоянием мозга. У большинства здоровых людей обнаруживается четкая корреляция между уровнем функциональной активности мозга в цикле «бодрствование — сон» и амплитудно-частотными характеристиками ЭЭГ. Снижение уровня бодрствования (переход от функционального покоя ко сну) сопровождается замедлением частоты и увеличением амплитуды волн ЭЭГ, а повышение уровня активности (переход от состояния покоя к психоэмоциональному напряжению) коррелирует с увеличением частоты активности (рис. 4).
Фазы сна
Сон здорового человека представляет собой циклическое повторение последовательно сменяющих друг друга фаз. Выделяют фазу медленного (ортодоксального) сна и фазу быстрого (парадоксального) сна. Медленный сон наступает сразу же после того, как человек заснул. Его продолжительность составляет 80—90 мин. Формирование и развитие медленного сна обеспечивается передними отделами гипоталамуса, ядрами шва, неспецифическими ядрами таламуса и средней частью моста (так называемым тормозным центром Моруцци). В структуре медленного сна выделяют три стадии. Стадии сна различаются по глубине сна, которую можно условно измерить по интенсивности внешнего стимула, способного вызвать пробуждение (рис. 5).
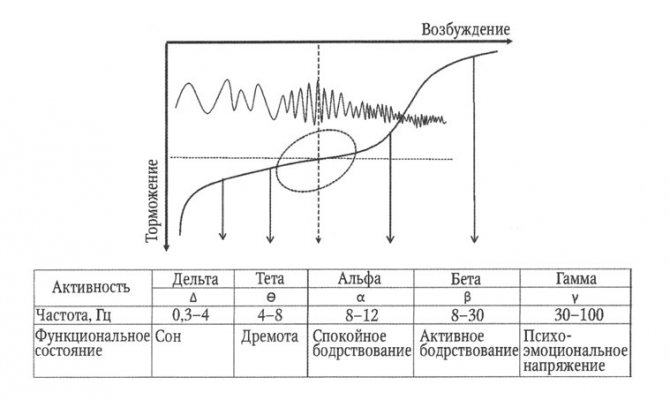
Рис. 4. Отражение в параметрах ЭЭГ основных нервных процессов: возбуждение и торможение.
На схеме альфа-ритм соответствует состоянию, при котором процессы возбуждения и торможения уравновешены по силе (зона, отмеченная эллипсом)
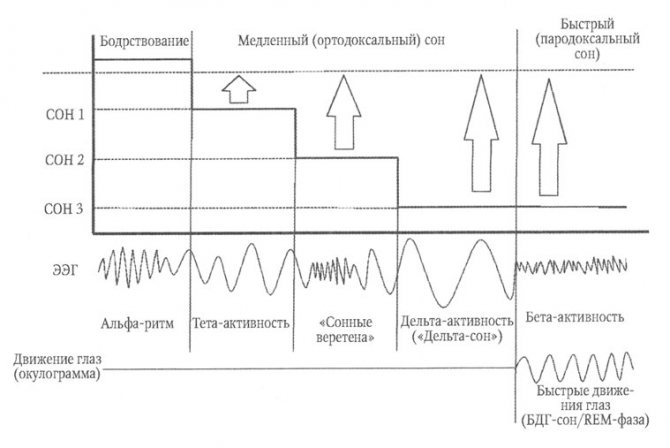
Рис. 5. Структура сна.
Стрелками схематично указана интенсивность стимула, способного вызвать пробуждение, как косвенное отражение глубины сна. Быстрый сон по глубине не уступает третьей стадии сна, что определяет парадоксальность состояния: глубокий сон на фоне быстроволновой активности на ЭЭГ и быстрые движения глазных яблок
Стадии сна
В первую стадию медленного сна человек пребывает в состоянии дремоты или полусна. Альфа-ритм замедляется, преобразуясь в низкоамплитудный тета-ритм. Активность мышц падает, частота пульса и дыхания снижается, обменные процессы замедляются, глазные яблоки совершают замедленное движение.
Во вторую стадию сна (легкий и неглубокий сон) на фоне преобладающей тета-активности появляются осцилляции низкоамплитудного сигма-ритма частотой 12—14—16 Гц, которые описываются как «сонные веретена». Вторая стадия сна по этой причине носит название «сон с веретенами». Вторая стадия — это примерно до 55 % от всего времени сна.
В третьей стадии медленного сна паттерн ЭЭГ представляет собой доминирование дельта-активности средней и высокой амплитуды. Третью стадию называют дельта-сном. Его глубина максимальна, разбудить человека в период дельта-сна крайне сложно. Третья стадия составляет около 10 % времени всего сна. Медленный сон обеспечивает полное восстановление затраченной за день энергии.
REM-sleep
В норме во время сна у человека эпизодически возникают периоды, во время которых наблюдается падение мышечного тонуса с одновременным появлением быстрых саккадических движений глазных яблок и иногда быстрых движений конечностей. В этот период спящего можно разбудить с помощью внешних стимулов интенсивностью не меньшей, чем в фазу глубокого сна. На ЭЭГ в течение этих периодов регистрируется полиморфная активность с преобладанием высоких частот. Исследования показали, что этим периодам соответствует субъективное переживание сновидения. Таким образом, эта стадия сна отличается рядом проявлений, которые противоречат друг другу: активация на ЭЭГ свидетельствует о переходе мозга на более высокий функциональный уровень, что противоречит большой глубине сна, измеряемой интенсивностью пробуждающего воздействия; высокий уровень субъективных эмоциональных и психических переживаний в сновидении противоречит мышечной релаксации. В связи с этим описанная фаза сна получила название парадоксальной, или «сон с быстрыми движениями глаз», сокращенно — БДГ-сон (англ. REM-sleep — rapid eye movement). Фазу парадоксального сна впервые выделил и подробно описал в 1953 г. Натаниел Клейтман совместно со своим учеником Юджином Асеринским.
Накопленная в электрофизиологии ЦНС фактология позволяет с достаточной надежностью связать каждый частотный диапазон ЭЭГ с определенным уровнем активности мозга в континууме «глубокий сон — психоэмоциональная напряженность». Альфа-ритм характерен для состояния функционального покоя. При переходе ко сну, что отражает развитие торможения, альфа-ритм заменяется на медленные ритмы, частота которых уменьшается пропорционально глубине сна, т. е. силе торможения. Повышение уровня бодрствования при умственной или физической деятельности сопровождается усилением возбудительного процесса, что сопровождается блокадой альфа-ритма и формированием высокочастотных ритмов, и чем выше уровень бодрствования, тем выше доминирующая частота на ЭЭГ.
Международный неврологический журнал 8 (46) 2011
Введение
Известно, что ни один из других видов электрических осцилляций мозга от 10–2 до 103 Гц не имеет такой функциональной значимости в процессах внимания, памяти, эмоций и мотивации, как веретенообразные осцилляции в диапазоне 5–15 Гц2 (Иваницкий, 1997; Ливанов 1989; Nunez et al., 2001). Показано, что функциональное ядро, организующее биоэлектрическую активность мозга в других частотных диапазонах, формируется осцилляциями, обладающими альфа-активностью (Martнnez-Montes et al., 2004; Klimesch et al., 2007). Однако до сих пор не выяснено, какие именно количественные признаки электроэнцефалограммы характеризуют флуктуации альфа-активности: увеличение или снижение частоты, рост или депрессия амплитуды, синхронность или десинхронизация фаз колебаний и в каком частотном диапазоне они проявляются. Кроме того, в связи с тем что не до конца ясно, какие нейрофизиологические механизмы лежат в основе феномена альфа-активности, трудно интерпретировать психологические корреляты ЭЭГ-признаков альфа-активности, тем более в некоем стандартизованном частотном диапазоне.
Цель настоящей работы — на основе анализа современных данных литературы и результатов собственных экспериментов определить, какова природа альфа-активности, какие признаки ЭЭГ ее характеризуют и каково их психофизиологическое значение.
Феномен «альфа-активность ЭЭГ»
В 1966 г. Грей Уолтер высказывал предположение, что существует большое разнообразие альфа-ритмов, которые отличаются по амплитуде в зависимости от локализации в коре, наличию или отсутствию блокады амплитуды при открывании глаз или при умственной нагрузке (Уолтер, 1966). Для частотных компонентов ЭЭГ такого типа употребляется выражение «семейство альфа-ритмов», куда включают: а) классический теменно-затылочный визуальный альфа-ритм (Adrian & Mathew, 1934; Kirschfeld, 2005); б) ритм, чувствительный к проприоцептивным раздражениям, называемый либо ролландическим (Зенков, 1996), либо сенсомоторным (Sterman, 1996 ), либо мю (Гусельников и Изнак,1983; Muthukumaraswamy et al., 2004), либо временно появляющимся «третьим» (Neidermayer,1993–1997), или лямбда-ритмом (Schomer, 2007); в) сонные веретено- образные волны (Ishii & Dziewas, 2001; Nunez et al., 2001). Кастро-Аламанкос и соавторы предпочитают не использовать термины «альфа-волны» или «сенсомоторный ритм», обозначая такие колебания как «приблизительно десятигерцевые осцилляции моторной коры» (Castro-Alamancos et al., 2007). Недавно гонконгскими исследователями Фонг была описана ритмическая альфа-подобная активность мозга в необычных диапазонах: при медленных (4–5 Гц) и быстрых (16–20 Гц) частотах ЭЭГ (Fong & Fong, 2001). Чапман и Лакелли называют ритм диапазона 4–12 Гц, зарегистрированный в гиппокампе, тета-ритмом (Chapman & Lacaille, 1999), а Моретти и соавторы тот же частотный диапазон, зарегистрированный в затылочных областях, называют альфа-ритмом (Moretti et al., 2004). Само существование семейства альфа-ритмов без четко определенных частотных и топографических границ приводит к предположению, что не только амплитуда и мощность осцилляций в неком стандартном диапазоне характеризуют альфа-активность, но и индивидуальное сочетание нескольких физических характеристик волнового процесса, имеющего четко определенную физиологическую значимость.
Изначально Ганс Бергер (1873–1941) определил альфа-активность как: 1) ритмический компонент ЭЭГ, частота которого лежит в интервале между 8 и 12 Гц; 2) колебания с наибольшей амплитудой в затылочной области коры, которые снижаются по амплитуде при открывании глаз и любых когнитивных нагрузках (Adrian & Matthews, 1934). В 1979 году группа чешских исследователей при анализе эффективности альфа-стимулирующего тренинга биоуправления установила, что доминирование по амплитуде в теменно-затылочной области может быть критерием для оценки альфа-активности только в случае, если параллельно оценивать длину альфа-веретена (Lansky & Bohdaneck, 1979). Теоретические расчеты (Осовец, 1983; Русинов и др., 1987; Hooper, 2005; Kirschfeld, 2005–2008; Tenke & Kayser, 2006) и анализ молекулярно-клеточных механизмов генерации альфа-осцилляций (Sherman & Guillery 2006; Steriade et al., 2001; Timofeev & Bazhenov, 2005) дали основание считать, что одной из наиболее информативных феноменологических характеристик альфа-активности является авторитмичность или веретенообразная организация этих волн (Ливанов, 1989; Каплан, 2005, Timofeev & Bazhenov, 2006).
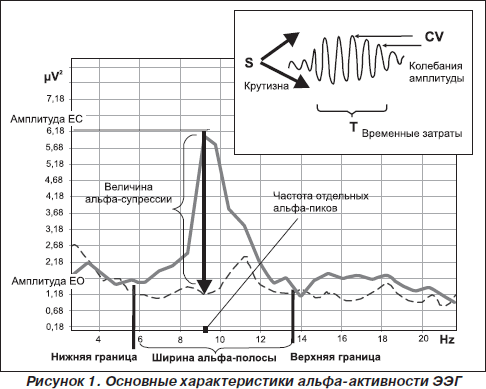
Таким образом, в современной литературе термин «альфа-ритм» применяется к любым компонентам ЭЭГ со следующими характеристиками: 1) частота доминирующего ритма ЭЭГ в теменно-затылочной области; 2) веретенообразность колебаний и 3) снижение амплитуды при реакции зрительной и когнитивной активации (Ливанов, 1989; Barry et al., 2007; Klimesh et al., 2007; Nunez et al., 2001; Thatcher et al., 2008) (рис. 1).
Природа альфа-осцилляций
Очевидно, что вышеперечисленные феноменологические характеристики альфа-активности обусловлены уникальной природой альфа-колебаний. Описание природы волнового процесса предполагает выяснение источника его генерации. В 1968 году П. Андерсен и С. Андерссон пришли к заключению, что ядра таламуса являются первичным генератором всех видов ритмической веретенообразной активности мозга (Andersen & Andersson, 1968). С тех пор исследователи добавили к гипотезе таламического источника генерации альфа-осцилляций только то, что в различных ядрах таламуса возникают веретенообразные электрические волны приблизительно раз в 100 мс, которые через таламокортикальные волокна формируют ритмическую активность соответствующих зон коры, так называемые кортикальные двойники (Andersen & Andersson, 1968; Jones et al., 2000; Contreras et al.,1997; Sherman S.M., Guillery R.W), и что эти волновые вспышки в каждой области коры контролируются определенным ядром таламуса (Hughes & Crunellia, 2007; Schreckenberger et al., 2004). Отличием таламических релейных клеток, в которых возникают эти волны, от других нейронов является повышенная плотность кальциевых T-каналов на мембранах поверхности клетки и эндоплазматической сети (Berridge et al., 2003; Sherman & Guillery, 2006). Установлено, что кальциевые каналы Т-типа, регулируя концентрацию кальция, могут тормозить передачу сигнала активации через таламус и тем самым стабилизировать состояние покоя (Page et al., 2006). Внутриклеточный кальциевый ток производит временную деполяризацию мембраны клетки с частотой приблизительно 10 раз в секунду, поскольку рефрактерность активации кальциевых каналов составляет приблизительно 100 мс (то есть это частота альфа-волн) Эти экспериментальные данные подтвердили гипотезу, выдвинутую М.Н. Ливановым в середине XX века и позднее лауреатом нобелевской премии Дж. Экклсом, о том, что организация ритмической активности мозга обусловлена рефрактерными свойствами возбуждения кальциевого тока, определяющими частоту ритмического разряда нейронов (Ливанов, Eccles, 1994). Таким образом, частота изменения концентрации кальция в цитоплазме нейрона является ключевым звеном изменения частоты альфа-активности мозга (Luthi et al., 1998).
1. Частота альфа-ритма. Наследуемость частоты доминантного ритма ЭЭГ и его межиндивидуально типологическая изменчивость доказаны многими исследованиями (Anokhin et al., 2006; Nunez et al., 2001; Smit et al., 2006; Thatcher et al., 2008). При этом среди разных показателей частоты альфа-активности (средней, средневзвешенной в альфа-диапазоне и частоты максимального пика) только индивидуальная частота максимального пика отражает единый агрегированный ресурс альфа-активности, а частоты других спектральных пиков из диапазона осцилляций, обладающих альфа-активностью, определяются интенсивностью стимула и длительностью его воздействия (Hooper, 2005). Грег Хупер теоретически рассчитал, что статистическое поведение величины индивидуальной частоты максимального альфа-пика в состоянии покоя в наибольшей степени соответствует законам нормального распределения по сравнению с показателями средней частоты и средневзвешенной в альфа-диапазоне (Hooper, 2005). Наши данные экспериментально подтверждают это положение (Базанова, 2010). Действительно, индивидуальная частота максимального спектрального пика альфа-ритма в задних отделах головного мозга в состоянии покоя с закрытыми глазами — показатель инвариантный, высоковоспроизводимый в тест-ретест-исследованиях и составляет 10,0 ± 0,5 Гц у большинства индивидов 25–40 лет (Bazanova, 2008a, Базанова, 2011; Kaiser, 2005; Nuwer, 2003; Thatcher et al., 2008). Наиболее вероятной причиной генетической детерминированности альфа-частоты является структурная организация кальциевых каналов, которая определяет частоту альфа-осцилляций (Anderson et al., 2005). Связь частоты генерации веретенообразных альфа-осцилляций со структурными носителями генетической информации — нуклеиновыми кислотами была продемонстрирована в 1965 году М.Б. Штарком (Штарк М.Б, 1965). Оказалось, что для лиц, отличающихся по показателю частоты максимального пика альфа-активности, характерны разные поведенческие стратегии (Базанова и Афтанас, 2006; Jin et al., 2006; Regehr & Kaplan, 1988). Однако наряду с этим установлена внутрииндивидуальная вариабельность частоты максимального пика альфа в зависимости от факторов возраста (Строганова и Цетлин, 1998; Bazanova, 2008a; Clarck et al., 2001), нейрогуморального статуса (Bazanova et al., 2008b; Creutzfeldt et al., 1976; Kamijo & Murakami, 2009; Korol, 2004; Tops et al., 2006), характера психофармакологического воздействия (Tops et al., 2006), выраженности процессов утомления (Kamei et al., 2000) (рис. 2 и 3). При этом беглость выполнения когнитивных (Doppelmayr et al., 2005; Klimesh et al., 2007; Moretti et al., 2007) и психомоторных заданий (Базанова и др., 2008; Hummel et al., 2004), а также степень владения профессиональными навыками (Базанова и др., 2008; Кураев и др., 2006) выше у лиц с высокой (> 10 Гц), чем с низкой (< 10 Гц) альфа-частотой. Вышеперечисленные внутрииндивидуальные флуктуации альфа-частоты обусловлены влиянием эпигенетических факторов, осуществляемым либо посредством воздействия на активность кальциевых каналов (Riegel & Williams, 2008), либо на процессы фосфорилирования протеинкиназ, участвующих в биосинтезе кальциевых каналов (Koulen et al., 2008), либо через действие на активность нейрональных рецепторов (Huang et al., 2007). При этом механизм формирования альфа-осцилляций, переданных посредством ионотропных рецепторов, иной, чем посредством метаботропных рецепторов (Pin & Duvoisin, 1995) — блокирование ионотропных рецепторов, через которые осуществляется генерация веретеноподобных волн, инициирует проведение осцилляций через метаботропные рецепторы (Lukatch et al., 2005). Метаботропные рецепторы, составляя 90–95 % синаптических входов, имеют проекцию по типу обратной связи в шестом слое коры и в стволовой ретикулярной формации (Pin & Duvoisin, 1995). Этим объясняется их ключевая роль в процессах торможения или так называемого нисходящего контроля (Klimesh et al., 2007). Так, недавними исследованиями П. Эмсона было продемонстрировано, что у нокаутных мышей по субъединицам метаботропных ГАМК-Б-рецепторов, как и ожидалось, нарушаются процессы их ингибиторного эффекта на активность кальциевых каналов, соответственно, отсутствует период рефрактерности, увеличивается частота и нарушается веретенообразность альфа-осцилляций (Emson, 2007).
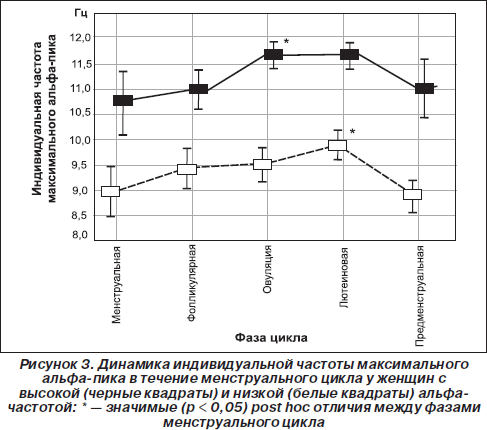
Интересно, что и сама частота нейрональных осцилляций может влиять на мембранную проводимость и чувствительность рецепторов к нейрогуморальным стимулам (Chi Ping et al., 2003; Destexhe & Sejnowski, 2003). Так, Чи Пинг и соавторы показали, что процесс эстрогеновой активации специфической Са2+-кальмодулинзависимой протеинкиназы, влияющей на частоту колебания концентрации кальция, линейно зависит от частоты входящих импульсов (Chi Ping et al., 2003). Возможно, поэтому повышение индивидуальной альфа-частоты при овуляции (когда происходит резкое увеличение концентрации эстрогенов) наблюдается только у женщин с высокой (≥ 10 Гц) альфа-частотой. А у женщин с низкой (< 10 Гц) альфа-частотой повышение частоты максимального пика происходит только на лютеиновой фазе (рис. 3), что наиболее вероятно связано с увеличением концентрации прогестерона (Baker et al., 2007; Bazanova et al., 2008b; Creutzfeldt et al., 1976). Другим примером того, что внутрииндивидуальные флуктуации альфа-частоты проявляются по-разному в зависимости от альфа-частотного паттерна ЭЭГ, является влияние возраста: снижение индивидуальной частоты альфа-пика после сорока лет происходит только у низкочастотных, но не наблюдается у высокочастотных испытуемых (Bazanova, 2008a). Это снижение тем более выражено, чем более ослабляются когнитивные способности (Сlarck et al., 2004) и увеличиваются показатели артериального давления (Bazanova, 2008a) (рис. 3).
Таким образом, в основе внутрииндивидуальной изменчивости частоты альфа-пика лежат механизмы молекулярной и структурно-рецепторной организации генерации и проведения импульса. В состоянии покоя индивидуальная частота максимального альфа-пика ЭЭГ отражает генетически детерминированные особенности структурной и молекулярной организации нейронов, а внутрииндивидуальная вариабельность этого показателя — изменения в кортикально-таламических и нейрогуморальных взаимодействиях, которые, в свою очередь, зависят от уровня индивидуальной частоты в состоянии покоя.
Возникает вопрос: отражаются ли генетически детерминированные различия по индивидуальной частоте максимального пика на других характеристиках альфа-активности?
2. Веретенообразность (авторитмичность) альфа-осцилляций отражает способность нейронных популяций к самонастройке или «модуляции фазы» — подстройке частоты осцилляций данного нейронального ансамбля к частоте другого, и тем самым координации их деятельности (Ливанов, 1989; Осовец, 1983). С.М. Осовец с соавторами (1983) на основании теоретических расчетов предположил, что взаимодействие ряда таких осцилляторов приводит к амплитудной и частотно-фазовой модуляции, поэтому веретена альфа-ритма носят нерегулярный характер и длительность их меняется в довольно широких пределах (Осовец, 1983; Kirschfeld, 2005; Лебедев и Луцкий, 1972; Каплан и др., 2002). При этом связи между отдельными осцилляторами внутри данного диапазона частот, очевидно, таковы, что система еще не входит в стохастический режим, но уже вышла из периодического. Такой процесс полностью соответствует современным представлениям о существовании промежуточных состояний при переходе от периодического к стохастическому процессу (или, на языке гидродинамики, — от ламинарного течения к турбулентному) (Осовец, 1983; Kirschfeld, 2005), что является следствием различной передачи импульсов, генерируемых по залповому или тоническому типу (Sherman & Guillery, 2006). Поскольку залпы или веретенообразные волны лучше распознаются корой и вызывают кортикальную активацию, их представляют как стимулы пробуждения — wake-up call коры, что может быть полезным для автоматически выполняемых действий (Steriade, 2001). Как только затрагиваются процессы внимания, начинает работать тонический запуск, для того чтобы максимизировать безукоризненную передачу информации по таламо-кортикально-таламическим цепям (Sherman & Guillery, 2006). На основании экспериментальных данных И. Тимофеев и М. Баженов пришли к заключению, что способность клеток продуцировать веретенообразные осцилляции зависит как от вида клеток, так и от условий, при которых эти осцилляции генерируются (Timofeev I., Bazhenov, 2005; Berridge et al., 2003).
До недавнего времени предполагалось, что сонные веретена и веретена альфа-осцилляций — совершенно разные феномены (Lopes da Silva, 1991, Steriade et al., 1990). Недавние работы, однако, показали, что залповый вид генерации колебаний отмечается не только во время сна, но и при бодрствовании ( Sherman & Guillery, 2006). C одной стороны, веретена ассоциируются с тормозящими процессами в таламусе и могут рассматриваться как входной механизм для блокирования процесса обработки информации из внешнего мира (Bazhenov et al., 2000; Berridge et al., 2003), а с другой, наоборот, выполняют роль зондирующего или контролирующего механизма связи организма с окружающей средой (например, веретенообразная электрическая активность мозга, отмечаемая с периодичностью раз в сутки у зимнеспящих животных (Штарк, 1970)).
Таким образом, наличие хорошо выраженных альфа-веретен на ЭЭГ свидетельствует об отлаженности адаптивных механизмов восходящего и нисходящего контроля (Giesbrecht et al., 2006), а их исчезновение сигнализирует о нарушениях механизмов контроля сенсомоторной интеграции у больных со спинальными травмами при отсутствии афферентации (Middleton et al., 2004) или при маниакально-депрессивном синдроме шизоидного типа (Каплан и др., 2005). Наши результаты свидетельствуют о том, что достижение успеха в тренинге оптимального психомоторного функционирования с помощью биоуправления соотносится с увеличением длительности и амплитуды альфа-веретена (Bazanova et al., 2008c). Как и ожидалось, вследствие различия формирования веретен при разных типах генерации и, соответственно, разной частоте импульсов мы отмечаем U-образную зависимость длительности веретенообразных сегментов от индивидуальной альфа-частоты. Самые короткие веретена отмечаются у лиц с крайне низкими и с крайне высокими значениями частоты альфа-пика (рис. 4). Эти же испытуемые со значениями индивидуальной частоты альфа-пика, наиболее отклоняющимися от среднего уровня, характеризуются наиболее высокими показателями оригинальности решения заданий в тесте Торренса (Базанова и Афтанас, 2007). Аналогичную инвертированную U-образную зависимость средней длины веретена альфа-ритма от его доминирующей частоты отметили А.Н. Лебедев и его сотрудники (Мальцева и Маслобоев, 1996). Предполагается, что более длительные альфа-веретена свидетельствуют о большей эмоциональной устойчивости и уравновешенности индивидов (Базанова и Афтанас, 2007, Лебедев и др., 2002). По данным А.Н. Лебедева, такие люди более дружелюбны и менее доминируют в межличностном общении (тест О. Лири), характеризуются большей гибкостью мышления, легкостью переключения с одного вида деятельности на другой и стремлением к разнообразию форм деятельности (Лебедев и др., 2002). Интересно, что у лиц с разной частотой альфа-активности в состоянии покоя с закрытыми глазами (то есть с различными молекулярно-генетическими механизмами генерации и передачи импульсов) веретна формируются и живут за счет различных механизмов, то есть одинаковая длительность альфа-веретена коррелирует с разными ЭЭГ-характеристиками (Bazanova et al., 2008c): у испытуемых низкочастотной группы длительность веретена тем больше, чем больше вариабельность амплитуды внутри веретена, а в высокочастотной группе — чем шире частотный диапазон, включаемый в реакцию активации. Что касается другой микроструктурной характеристики веретена — вариабельности амплитуды, которая свидетельствует о более интенсивном процессе фазовых перестроек (Oprisan et al., 2004), было обнаружено, что наиболее высокий коэффициент вариабельности и более широкий альфа-частотный диапазон на энцефалограмме покоя отмечается у лиц, характеризующихся более высокими коэффициентами невербальной креативности Торренса (Базанова и Афтанас, 2007–2008), а также у музыкантов, обладающих наиболее высоким уровнем исполнительского мастерства (Bazanova et al., 2003).
Таким образом, важнейшей особенностью альфа-осцилляций является их веретенообразность, или высокая фазическая изменчивость, отражающая динамику ансамблевой организации корковой нейронной активности. При этом 1) длительность веретена соотносится со временем жизни нейронного ансамбля, что может служить маркером когнитивной работоспособности, эмоциональной устойчивости; 2) внутрисегментная амплитудная вариативность или интенсивность фазовой перестройки в рамках ансамбля соответствует поиску оптимальных условий контроля процессов восходящей и нисходящей информации и свидетельствует об уровне креативности; 3) крутизна межсегментных переходов позволяет судить о скорости формирования или распада нейронных ансамблей.
Наконец, третья функциональная особенность альфа-осцилляций, которая была описана еще в 1934 г. Е. Эдриан и Б. Матьюс, — это снижение амплитуды альфа-волн в ответ на открывание глаз, названное Berger effect (Adtian & Matthew, 1934). Роберт Барри предложил для обозначения реактивности альфа-ритма на зрительную стимуляцию использовать термин «активация» (Barry et al., 2007). Глубина снижения альфа-амплитуды ЭЭГ принимается за интенсивность активации (Barry et al., 2007) (рис. 1). Установлено, что сила реакции активации меньше у испытуемых в состоянии покоя по сравнению с условиями, при которых необходим процесс внутренней переработки информации (Cooper et al., 2006). Поскольку было показано, что длительная зрительная стимуляция не только вызывает потенциал действия, но и снижает чувствительность нейронов к свету (Kirschfeld, 2008; Lansky et al., 1979), длительность супрессии альфа-амплитуды при открывании глаз является одним из информативных признаков стабильности реакции активации и вместе с глубиной десинхронизации соотносится с эффективностью интеллектуальной деятельности (Shmelkina, 1999; Stipacek et al., 2003). Неудивительно, что ряд исследователей используют показатели супрессии амплитуды в ответ на открывание глаз в качестве индивидуально-типологических признаков, имеющих отношение как к общей адаптивности (Arikan et al., 2006; Bazanova et al., 2003), так и к формированию изменений когнитивной деятельности (Alexander et al., 2006; Stipacek et al., 2003) и успешности в обучении с использованием адаптивной обратной связи (Базанова и др., 2007; Shmelkina, 1999). Так же, как другие характеристики альфа-активности ЭЭГ, процесс активации, с одной стороны, в значительной мере определяется генетическими факторами (Малых и др., 1998; Anokhin et al., 2006), а с другой — варьирует в зависимости от возраста (Andersonr et al., 2005; Bazanova et al., 2008а), нейрогуморального статуса (Bazanova et al., 2008b; Goldstein et al., 2005) и частоты доминирующих альфа-осцилляций (Gebber et al., 1999; Mazaheri and Jensen, 2006). Однако утверждение, что амплитуда альфа-ритма снижается только вследствие стимуляции светом, становится сомнительным, когда открывание глаз в абсолютно темной комнате также сопровождается снижением амплитуды альфа-осцилляций, и не только в зрительной коре (Klimesch et al., 2001; Toscani et al., 2010). Согласно данным М. Мусманн и соавторов, снижение альфа-амплитуды в темной комнате происходит не вследствие первичной сенсорной стимуляции, а, что более вероятно, отражает нисходящее контролирующее влияние коры (Moosmann et al., 2003). Депрессия амплитуды линейно зависит от активности рецепторов, величины стимула и, соответственно, типа генерации (Sherman & Guillery, 2006). Поэтому ЭЭГ-ответы на зрительную стимуляцию могут быть как аддитивными, так и являться результатом фазовой перестройки (David et al., 2005; Kirschfeld, 2005, 2008). Эти различные механизмы активации мы наблюдаем у лиц с разной частотой альфа в состоянии покоя. Множественный регрессионный анализ показал, что сила реакции зрительной активации у низкочастотных испытуемых была тем больше, чем больше амплитудная вариативность альфа-веретена (то есть интенсивность фазовых перестроек), а у высокочастотных — чем выше амплитуда альфа-осцилляций (Bazanova et al., 2008c).
На основании вышесказанного можно предположить, что изучение реакции на зрительную стимуляцию позволяет выявить индивидуальные частотные компоненты ЭЭГ, которые обладают альфа-активностью и которые, соответственно, можно назвать альфа-ритмом. Однако до настоящего времени определение частотных диапазонов проводится согласно договоренностям и стандартам (Kaiser, 2004), не учитывающим индивидуальные проявления реакции снижения амплитуды в ответ на зрительную стимуляцию. Между тем в недавних пионерских исследованиях в условиях сочетанной регистрации ЭЭГ и ф-МРТ убедительно показано, что эпизоды спонтанного снижения альфа-мощности ЭЭГ в состоянии физиологического покоя ассоциируются с усилением мозговой активности (Laufs et al., 2006). Авторы этого и других аналогичных исследований предполагают, что картина мозговой активации во время снижения альфа-мощности зависит от фонового уровня мозговой активности, наблюдаемой в более широком (не только в стандартном альфа) спектральном диапазоне ЭЭГ (Jones et al., 2000; Kaiser, 2005). Результатами наших исследований также продемонстрировано, что индивидуальная ширина альфа-диапазона варьирует на меж- и внутрииндивидуальном уровне и зависит от многих факторов: возраста, нейрогуморального статуса, уровня профессионального мастерства и пр. (Bazanova et al., 2008 a, b, c). Поскольку границы тета- и бета-частотных диапазонов ЭЭГ будут определяться в зависимости от границ альфа-полосы, учет значений индивидуальной ширины альфа-диапазона становится особенно актуальным при проведении тренингов нейробиоуправления, организованных при использовании мощности не только альфа-, но и тета- и бета-диапазонов ЭЭГ в качестве сигналов обратной связи (Базанова и Афтанас, 2007; Kaiser, 2005).
Таким образом, характеристики реактивности на зрительную стимуляцию — глубина и длительность снижения мощности альфа-осцилляций и ширина диапазона, в котором отмечается реакция активации, могут служить индивидуальными маркерами альфа-активности мозга. Так же, как длительность альфа-веретена, сила активации не отличается по средним значениям у испытуемых с различной индивидуальной частотой альфа-осцилляций, но она обусловлена различными механизмами.
Представленные выше нейрофизиологические механизмы альфа-активности и их взаимосвязь с когнитивными и физиологическими функциями дают основание считать, что для количественной оценки альфа-активности мозга необходимо выяснить как минимум величину трех основных ЭЭГ-признаков:
1) амплитуды и частоты доминирующих осцилляций в теменно-затылочной области в состоянии покоя с закрытыми глазами;
2) авторитмичности — по характеристикам микроструктуры альфа-веретена: внутрисегментной вариабельности амплитуды, длительности, средней амплитуде и крутизне нарастания веретена;
3) зрительной активации по показателям глубины и стабильности снижения амплитуды, а также по ширине частотного диапазона, в котором происходит активация.
Очевидно, что измерение какой-либо отдельной характеристики, например частоты или амплитуды, не может претендовать на полноту оценки альфа-активности, а анализ вместе взятых индивидуальных параметров альфа-активности ЭЭГ и их взаимосвязей может привести к выяснению особенностей работы психофизиологических механизмов поведения и адаптации.
Половые различия ЭЭГ
Существуют определенные половые различия ЭЭГ. Женщинам свойственны более высокие частоты a-ритма и большее количество β-активности. Эти данные определенным образом коррелируют с половыми различиями психологических характеристик, свидетельствуют о более высоком уровне активации у женщин, очевидно, имеют генетическую обусловленность и могут быть связаны с гормональными особенностями4. Показано, что в предменструальный период наблюдается увеличение частоты a-ритма, коррелирующее с повышением психометрических показателей. В этот же период увеличивается уровень прогестерона. У женщин, принимающих гормональные контрацептивы, не обнаруживают такой цикличности на ЭЭГ, они отличаются более низкими психометрическими показателями5.
Что можно оценить при ЭЭГ?
Данный вид обследования позволяет определить функционирование отделов головного мозга при разных состояниях организма – сне, бодрствовании, активной физической, умственной деятельности и других. ЭЭГ – это простой, абсолютно безвредный и безопасный метод, не нуждающийся в нарушении кожных покровов и слизистой оболочки органа.
В настоящее время он широко востребован в неврологической практике, поскольку дает возможность диагностировать эпилепсию, с высокой степенью выявлять воспалительные, дегенеративные и сосудистые нарушения в мозговых отделах. Также процедура обеспечивает определение конкретного месторасположения новообразований, кистозных разрастаний и структурных повреждений в результате травмы.
ЭЭГ с применением световых и звуковых раздражителей позволяет отличить истерические патологии от истинных, или выявить симуляцию последних. Процедура стала практически незаменимой для реанимационных палат, обеспечивая динамическое наблюдение коматозных пациентов.
Пропадание на ЭЭГ сигналов эклектической активности свидетельствует о наступлении летального исхода
Редкие типы ЭЭГ
С точки зрения диагностики, представляют интерес некоторые относительно редкие типы ЭЭГ. У части здоровых обследуемых отсутствует нормальный а-ритм, и вместо него регистрируется активность 14-18 Гц. Эта активность имеет максимальную амплитуду порядка 50 мкВ в затылочных отделах и, подобно нормальному а-ритму, ее амплитуда снижается по направлению кпереди. В ответ на афферентные стимулы при умственной нагрузке, эмоциональном напряжении этот ритм исчезает и возникает реакция активации. Все это позволяет рассматривать указанную активность как эквивалент a-ритма и расценивать такие ЭЭГ как вариант нормы (рис. 6).
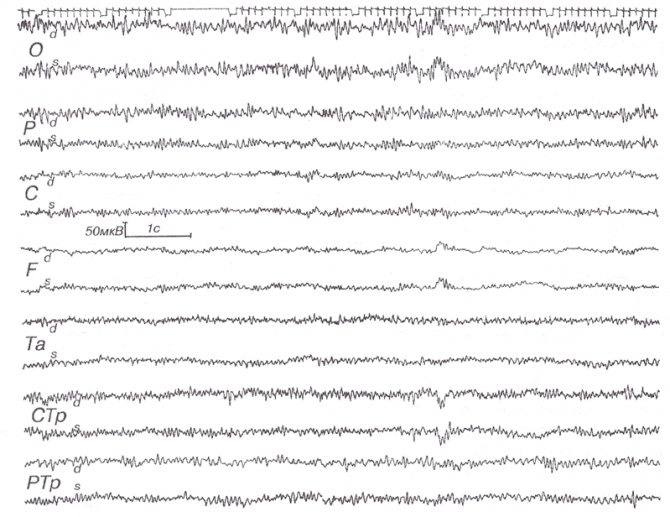
Рис. 6. Быстрый a-вариант.
Неврологически здоровый В.Н.К., 43 лет. Регулярный ритм 13-17 Гц при закрытых глазах с амплитудным градиентом, соответствующим нормальному а-ритму.
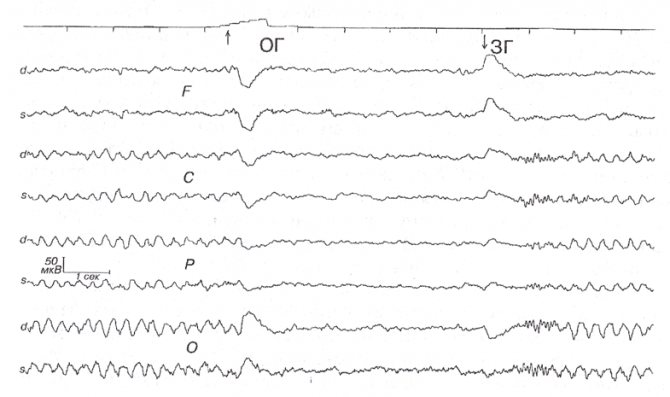
Рис. 7. Медленный а-вариант.
Женщина Ф.Е.А., 27 лет, с психогенными головными болями без органической неврологической патологии. Регулярный ритм 3,6 Гц при закрытых глазах с амплитудным градиентом, соответствующим нормальному a-ритму. Открывание глаз («ОГ») вызывает реакцию десинхронизации. При закрывании глаз («ЗГ») после короткого веретена а-активности 13-14 Гц — перестройка на «медленный a-вариант». Видны характерные артефакты движений глаз при их закрывании и открывании.
Наконец, очень редко (около 0,2% случаев, по данным I.Petersen, R.Sorbie, 1962) встречаются своеобразные ЭЭГ, на которых при закрытых глазах в затылочных отделах регистрируются регулярные, близкие к синусоидальным, медленные волны с частотой 2,5-4 Гц и амплитудой 50-80 мкВ. Этот ритм исчезает при афферентных стимулах с появлением типичной реакции, как и a-ритм. При открытых глазах на ЭЭГ регистрируется нормальная низкоамплитудная полиморфная активность в диапазоне а- и β-ритма. Клинически в этих случаях не удается выявить органической церебральной патологии, и жалобы носят невротический, функциональный характер. В нашем собственном наблюдении больная, помимо головных болей, расцененных в результате полного клинического обследования как психогенные, жаловалась на хроническую бессонницу (рис. 7). Не исключено поэтому, что подобного типа ЭЭГ могла служить проявлением нарушения нормальной функции регуляции сна и бодрствования. Такого рода ЭЭГ следует рассматривать как пограничные между нормой и патологией; по мнению I.Peterson, R.Sorbie6, они могут указывать на дисфункцию диэнцефальных неспецифических систем мозга.
О ЧЁМ МОЖНО УЗНАТЬ ИЗ ЭЭГ
Медицинская статистика и опыт специалиста позволяют расшифровать аномалии на графиках ЭЭГ и диагностировать их причины.
Аномальные характеристики ЭЭГ, указывающие на всевозможные патологии:
- Сигналы от нейронов полушарий не согласованы и несимметричны;
- Основные ритмы имеют резкие частотные перепады, соответствующие резким всплескам активности или внезапному их спаду. Наличие скачков частот указывают на присутствие инфекции, опухолей, травматических повреждений или на развитие инсульта.
- Чередование заостренных и базальных петель, большой разбег частот, одноразовые всплески или серийные могут означать эпилепсию. Но между приступами показатели ЭЭГ могут быть как у небольного человека.
- Появление δ- и θ-ритмов у неспящего пациента указывает на аномалии мозга.
Нарушения основных ритмов и патологии:
- асимметрия α-ритма в полушариях до 30% свидетельствует об опухолях или инфаркте/инсульте. Иногда α-петли имеют высокие частотные значения, сопровождающиеся некоторой нестабильностью, что показывает наличие поражений при черепно-мозговой травме. При старческом слабоумии или возникшем из-за травм α-волны исчезают или становятся асимметричными. Отклонения α-ритма у детей может быть вызвана задержкой развития психомоторных навыков.
- β-петли с амплитудой 50 мкВ и более свидетельствуют о сотрясения мозга. Короткие всплески β-волн указывают на энцефалит. Их нарастающая частота и длительность – признак воспалительного процесса.
- Амплитуда δ-волн более 40 мкВ – признак аномалий мозговой деятельности. Когда это проявляется во всех участках, то причиной являются сложные болезни ЦНС. При новообразованиях δ-ритм также характеризуется большими разбросами колебаний.
- Явно выраженные θ- и δ-пики в затылочной части ЭЭГ головного мозга у детей свидетельствуют о задержке развития.
Такие признаки, как отсутствие острых пиков, синхронность и симметрия ритмов в полушариях, преобладание α- и β-ритмов у неспящих, стабильность сигналов мозга при кратковременных световых воздействиях, свидетельствуют о нормальной деятельности мозга здорового человека.
ЭЭГ КОМАТОЗНОГО СОСТОЯНИЯ
Если деятельность ЦНС нивелирована сильнодействующими лекарствами или нарушено кровоснабжение мозга, на ЭЭГ будет фиксироваться частичное или полное отсутствие активности. Фактически в этом состоянии жизнедеятельность организма поддерживается искусственно.
Провоцирующие пробы (процедуры активации)
➥ Основная статья: Функциональные пробы ЭЭГ
Провоцирующие пробы (или процедуры активации) служат важным компонентом ЭЭГ в клинической практике и представляют собой различные типы раздражителей или воздействий, которые способны провоцировать нарушения на ЭЭГ. Традиционной практикой является проведение гипервентиляции и ритмической фотостимуляции (эти пробы провоцируют появление замедления и/или эпилептиформных нарушений, хотя возможно применение и других провоцирующих проб — депривация сна, фармакологические и другие методы.
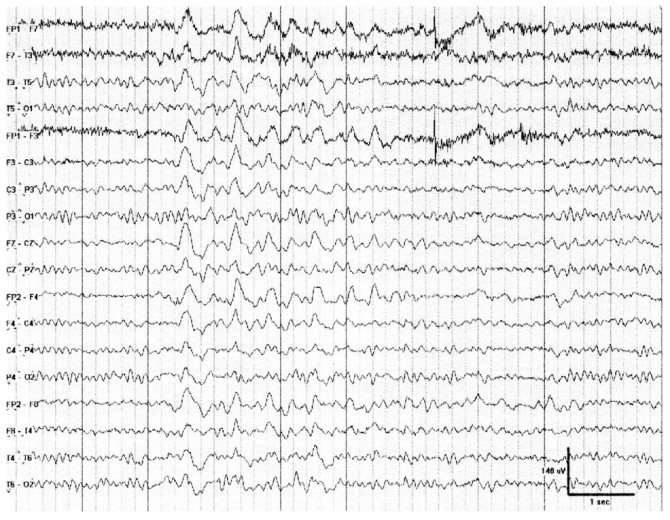
Рис. 13. Нормальное увеличение амплитуды ЭЭГ («накопление» — build-up) в процессе гипервентиляции
Гипервентиляция обычно проводится в течение 3—5 мин в большинстве ЭЭГ-лабораторий. Цель — достижение церебральной вазоконстрикции с помощью глубокого дыхания, в результате развития системной гипокарбии. Гипервентиляция в норме вызывает билатеральное усиление активности θ- и δ-частот («накопление» — build-up7) с преобладанием в лобных отведениях, часто имеющей высокую амплитуду. Прекращение данного эффекта наблюдается в норме в течение 1 мин. У пациентов с локализационно-обусловленной эпилепсией активация или генерация эпилептиформных разрядов встречается не часто (< 10%); однако этот показатель может достигать 80% у пациентов с генерализованными эпилепсиями, включая абсансные приступы. Гипервентиляция может провоцировать фокальное замедление у пациентов с лежащим в основе структурным поражением. Гипервентиляцию не следует проводить у пациентов с тяжелыми сердечными или легочными заболеваниями, при остром или недавно перенесенном инсульте, серьезных сердечно-сосудистых заболеваниях с поражением крупных сосудов, при серповидноклеточной анемии или серповидноклеточной аномалии эритроцитов. Также эту пробу следует использовать с осторожностью во время беременности.
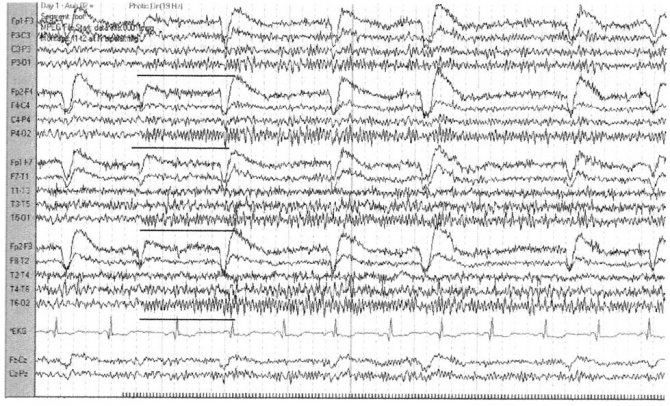
Рис. 14. Усвоение ритма на частоте 20 Гц
регистрируемое в отведениях Р3-О1, Р4-О2, Т5-О1 и Т6-О2
Ритмическая фотостимуляция в норме вызывает возникновение потенциалов, четко связанных со временем появления и частотой прерывистых световых вспышек, это явление обозначают как усвоение ритма. Ответ зависит от фонового освещения и расстояния от источника света до пациента. С целью оптимизации эффекта стимуляции выбирают расстояния <30 см от пациента. Вспышки света очень короткие, и световая стимуляция проводится последовательно на частотах от 1 до 30 Гц, действие каждого стимула продолжается приблизительно 10 с. Возможно определение субгармонических и гармонических частот колебаний при световой стимуляции. Усвоение ритма обычно наблюдается в затылочных отведениях, на частотах, приближающихся к частоте α-ритма, при закрытых глазах. Фотомиоклонический (или фотомиогенный) ответ представляет собой доминирующие в лобных отведениях мышечные артефакты, которые возникают, если вспышки света провоцируют повторяющиеся локальные сокращения лобных мышц (фотомиогенный ответ). Возможно также вовлечение периокулярных мышц с появлением одиночных вызванных световым мельканием молниеносных подергиваний головы (фотомиоклонический ответ). Миогенные спайки появляются через 50—60 мс после вспышки и увеличиваются по амплитуде по мере повышения частоты стимуляции. Данный ответ считается нормальным, хотя может наблюдаться при синдроме отмены (абстиненции) или при состояниях гипервозбудимости.
Процесс изучения результатов
Анализ полученных результатов проводится параллельно во время процедуры, и в ходе фиксации показателей, и продолжается по ее окончании. При записи учитываются присутствие артефактов – механического движения электродов, электрокардиограммы, электромиограммы, наведение полей сетевого тока. Оценивается амплитуда и частота, выделяют наиболее характерные графические элементы, определяют их временное и пространственное распределение.
По окончании производится пато- и физиологическая интерпретация материалов, и на ее базе формулируется заключение ЭЭГ. По окончании заполняется основной медицинский формуляр по данной процедуре, имеющий название «клинико-электроэнцефалографическое заключение», составленный диагностом на проанализированных данных «сырой» записи.
Расшифровка заключения ЭЭГ формируется на базе свода правил и состоит из трех разделов:
- Описание ведущих видов активности и графических элементов.
- Вывод после описания с интерпретированными патофизиологическими материалами.
- Корреляция показателей двух первых частей с клиническими материалами.
Основным описательным термином в ЭЭГ является «активность», он оценивает любую очередность волн (активность острых волн, альфа-активность и др.).
Список дополнительной литературы
- Александров М. В., Иванов Л. Б., Лытаев С. А. [и др.]. Электроэнцефалография : руководство / под ред. М. В. Александрова. — 3-е изд., перераб. и доп. — СПб.: СпецЛит, 2021. — 224 с.
- Abou-Knalil В., Misulis К.Е. Atlas of EEG and Seizure Semiology. Butterworth Heinemann, Philadelphia, 2006: 1-213.
- Benbadis S.R., Tatum W.O. Overinterpretation of EEGs and misdiagnosis of epilepsy. J. Clin. Neurophysiol. 2003; 20: 42-44.
- Blume W.T., Masako K., Young G.B. Atlas of Adult Electroencephalography. 2nd ed. Lippincott Williams & Wilkins, Philadelphia, 2002: 1—531.
- Kellaway Peter. Orderly approach to visual analysis: elements of the normal EEG and their characteristics in children and adults. In: Ebersole J.S., Pedley T.A., eds. Current Practice of Clinical Electroencephalography. 3rd ed. Lippincott Williams & Wilkins, Philadelphia, 2003: 100—159.
- Markand, Omkar N. Pearls, perils, and pitfalls in the use of the electroencephalogram. Semin. Neurol. 2003; 23 (1): 7-46.
- Olejniczak P. Neurophysiologic basis of EEG. J. Clin. Neurophysiol. 2006; 23 (3): 186-189.
- Tatum W.O. IV, Husain A., Benbadis S.R., Kaplan P.W. Normal human adult EEG and normal variants. J. Clin. Neurophysiol. 2006; 23 (3): 194-207.
- Westmoreland B.E Benign electroencephalographic variants and patterns of uncertain clinical significance. In: Ebersole J.S., Pedley T.A., eds. Current Practice of Clinical Electroencephalography. 3rd ed. Lippincott Williams & Wilkins, Philadelphia, 2003: 235—245.
Footnotes
- Adams A.E. Frequenzanalyse des flachen EEG I Dtsch. Z. Nervenheilk., 1968, Bd. 193., s. 57-72.
- Небылицын В.Д. Основные свойства нервной системы человека. — М.: Изд-во АПН РСФСР, 1966, 386 с.
- Saul L.J. Davis H., Davis I’.A. Psychologic correlations with the electroencephalogram / Psychosom. med., 1949, v. 11, p. 361—376.
- Friedl W., Vogel F. Geschlechtsunterschiede im normalen Ruhe-EEG bei jungen Erwachsenen / Z. EEG-EMG. 1979, Bd. 10, s. 70-79.
- Creutzfeldt O.D., Arnold P.M., Becker D. et al. EEG changes during spontaneous and controlled menstrual cycles and their correlation with psychological performance I Electroenceph. clin Neurophysiol. 1976, v. 40, №2, p. 113—131.
- Petersen I., Sorbie R. Slow posterior rhythm in adults / Electroenceph. clin. Neurophysiol., 1962, v. 14, p. 161 — 170.
- Термин используется для описания нарастающего увеличения вольтажа ЭЭГ или появления волн с нарастающей амплитудой, во время гипервентиляции часто сопровождающейся снижением частоты колебаний. Иногда применяется в описании гипервентиляции или разрядов приступа. Использование термина не рекомендуется
- С. Гуляев, И. Архипенко, А. Овчинникова. Электроэнцефалография в практике врача невролога. Saarbrücken: LAP, 2013. С. 64.
- С. Гуляев, И. Архипенко, А. Овчинникова Электроэнцефалография в практике врача невролога. Saarbrücken: LAP, 2013. С. 64.
- С. Гуляев, И. Архипенко, А. Овчинникова Электроэнцефалография в практике врача невролога. Saarbrücken: LAP, 2013. С. 65.
- С. Гуляев, И. Архипенко, А. Овчинникова Электроэнцефалография в практике врача невролога. Saarbrücken: LAP, 2013. С. 65.
- С. Гуляев, И. Архипенко, А. Овчинникова Электроэнцефалография в практике врача невролога. Saarbrücken: LAP, 2013. С. 66.