16 900
30 000
Интимная пластика со скидкой 43%
«Медлайн-Сервис» предлагает провести интимную пластику больших или малых половых губ за 16 900 руб. вместо 30 000 руб.
Условия и цены
17 000
26 440
Программа «Счастливые родители» со скидкой 35%
«Медлайн-Сервис» предлагает программу «Счастливые родители» со скидкой 35%
Условия и цены
95 000
118 970
Программа дородового наблюдения «Стандарт +»
«Медлайн-Сервис» предлагает программу дородового наблюдения «Стандарт +» (ведение беременности) за 95 000 руб. вместо 118 970 руб.
Условия и цены
66 000
83 180
Программа дородового наблюдения «Стандарт»
«Медлайн-Сервис» предлагает программу дородового наблюдения «Стандарт» (ведение беременности) за 66 000 руб. вместо 83 180 руб.
Условия и цены
5 500
8 820
Экспресс-диагностика – Check-UP «Женское здоровье» со скидкой 35%
«Медлайн-Сервис» предлагает экспресс-диагностику – Check-UP «Женское здоровье» со скидкой 35%
Условия и цены
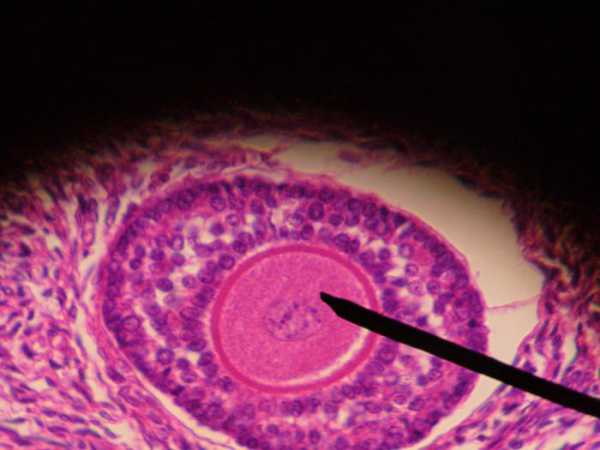
Фолликулы – это структурные составляющие половых желез у женщин. Один из этих элементов, называемый доминантным, во время овуляции освобождает созревшую для зачатия яйцеклетку. При нормальном строении фолликула, а также его своевременном дозревании, женщина способна забеременеть. Любые отклонения от нормы могут привести к развитию кисты на яичнике или даже бесплодию. Существует множество причин, способных вызвать данные нарушения, поэтому при возникновении любых необычных симптомов следует незамедлительно обратиться за медицинской помощью.
Функции и назначение фолликулов
Фолликул – это недозревшая яйцеклетка, которая окружена слоем эпителиальных клеток и двойным покровом соединительной ткани. Основной ее задачей является защита половой клетки от негативного воздействия различных факторов. Именно от данных элементов зависит правильное дозревание яйцеклетки и ее оплодотворение, и, следовательно, возможность забеременеть и вынашивать ребенка. Также они предназначены для выработки женского гормона – эстрогена.
У женщин половая система и фолликулярный аппарат развиваются в перинатальном периоде, и на данном этапе устанавливается постоянное количество фолликулов, которое остается неизменным на протяжении всей жизни (30 – 50 тысяч).
Различают несколько этапов развития фолликулов в яичниках:
- Образование нескольких мелких недозревших клеток.
- На 5 день УЗИ позволяет увидеть до десяти антральных фолликулов на периферии яичника (их размер колеблется в пределах 2 – 4 мм).
- По истечении семи дней их размер увеличивается до 6 мм, что дает возможность рассмотреть сеть капилляров на их основании.
- На восьмой день определяются доминантные фолликулы, которые продолжают расти и развиваться.
- Примерно на 10 день УЗИ дает возможность выявить доминантный фолликул, имеющий самый большой размер (около 15 мм), в то время как остальные будут вдвое меньше.
- Спустя две недели доминант достигает размеров 25 мм. В этот момент происходит активная выработка женского гормона – эстрогена, под воздействием которого защитная оболочка прорывается (позволяя яйцеклетке выйти наружу) и на 15 – 16 день происходит овуляция.
- Яйцеклетка попадает в маточные трубы. В результате ее встречи со сперматозоидом происходит оплодотворение. В противном случае во время месячных она выходит из матки вместе с эпителием.
Созревание в каждом менструальном цикле
С начала менструального цикла в обоих яичниках образуется порядка 8-10 вторичных фолликулов. Примерно с восьмого или девятого дня цикла пузырьки начинают заполняться образующейся под воздействием синтезируемых женским организмом эстрогенов жидкостью. И уже на данном этапе заметен доминантный фолликул: он больше остальных, и это можно увидеть на УЗИ.
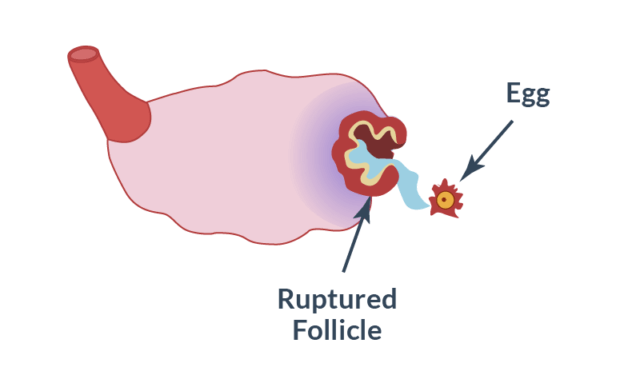
Выход яйцеклетки из фолликула.
Пузырёк продолжает заполняться жидкостью, растягивается и в момент овуляции лопается. Выходит созревшая яйцеклетка, которая начнёт продвигаться по фаллопиевой трубе в матку, чтобы соединиться со сперматозоидом. На какой день происходит разрыв? Это зависит от продолжительности менструального цикла: если он длится 28-30 дней, то овуляция и, соответственно, выход яйцеклетки из лопнувшего фолликула, приходится на 14-16-ый день (отсчёт ведётся с начала менструации).
На месте разорвавшегося пузырька образуется жёлтое тело – временная железа внутренней секреции, активно синтезирующая прогестерон и обеспечивающая подготовку матки к возможной беременности. Вырабатываемый гормон делает эндометрий рыхлым и мягким, чтобы плодное яйцо смогло прочно в нём закрепиться и начать развиваться.
Нарушения нормы
Какова же норма фолликулов в яичнике? Избыток или недостаток данных элементов является нарушением. Если в одном яичнике находится более десяти недозревших яйцеклеток и эта цифра остается неизменной на протяжении всего цикла, можно говорить о превышении нормы (выявить это позволяет исключительно УЗИ). В зависимости от количества элементов, обнаруженных во время ультразвукового исследования, можно сделать следующее заключение:
- в диапазоне от семи до шестнадцати – фолликулов в яичнике много и вероятность зачатия высокая;
- от четырех до шести – небольшая вероятность беременности;
- менее четырех – вероятность зачатия практически отсутствует.
Однако, это не всегда свидетельствует о какой-либо патологии и может быть вызвано стрессами, переживаниями или переутомлением. В этом случае количество фолликулов в яичниках нормализуется после первой овуляции. Терапия проводится, если нарушения вызваны следующими причинами:
- неправильным выбором контрацептивов;
- нарушением функционирования щитовидной железы;
- быстрым набором или потерей веса;
- сбоем в работе эндокринной системы;
- повышенным содержанием пролактина в организме.
Выявить вышеперечисленные нарушения можно при помощи диагностических процедур.
Недостаточное количество или отсутствие половых клеток может быть вызвано гормональным сбоем или ранним климаксом. Обычно данную проблему отслеживают на седьмой день менструального цикла. В этом случае для лечения репродуктивной функции женщины назначаются гормональные препараты.
Подводя итоги, можно выделить два существующих варианта развития фолликула:
- Протекание менструального цикла с одним доминантом в левом или правом яичнике.
- Его отсутствие, в результате — яйцеклетка не вызревает, происходит нарушение цикла менструации. В этом случае зачатие невозможно.
Возможные патологии
Рассмотрим некоторые отклонения от нормы:
- Доминантный фолликул отсутствует. Это говорит о том, что овуляции в текущем менструальном цикле, скорее всего, не будет. Ановуляторные циклы бывают у каждой здоровой женщины один или два раза в год. Если овуляция отсутствует несколько месяцев подряд, это ненормально.
- Множественные фолликулы или так называемые мультифолликулярные яичники – это отклонение, развивающееся в результате гормональных нарушений. Доминантный фолликул может отсутствовать или развиваться медленно, что снизит вероятность зачатия.
- Образование кист. Доминантный фолликул не лопается, переполняется жидкостью и растягивается, формируя доброкачественное образование – кисту (она может расти или самостоятельно регрессировать, то есть лопнуть и исчезнуть).
- Атрезия – замедление, остановка роста главного пузырька и его последующее отмирание без выхода созревшей яйцеклетки.
- Персистенция. Доминантный фолликул достигает нужных размеров, но не разрывается и сохраняется в неизменном целом виде до начала менструации. Зачатие становится невозможным.
- Лютеинизация. Жёлтое тело начинает формироваться при наличии в яичнике целого фолликула.
Перечисленные патологии заметны на УЗИ и обусловливаются гормональными сбоями или заболеваниями репродуктивной системы.
Доминантный фолликул необходим для оплодотворения. Но зачатие произойдёт, если пузырёк правильно сформируется, и из него выйдет созревшая яйцеклетка. Изложенная в статье информация поможет разобраться в механизме оплодотворения и выявить некоторые проблемы.
( проголосовало: 5, оценка: 4,80 из 5)
Поделиться новостью в соцсетях
Задать вопрос! У вас есть вопросы? Не стесняйтесь, задавайте любые! И наш штатный специалист поможет вам. Перейти>>
Метки: фолликул
- Рекомендуемые статьи
- Планирование беременности при хроническом эндометрите
- Опасные дни для зачатия
- Эрозия шейки матки при планировании беременности
« Предыдущая запись
Доминантный фолликул
В середине цикла обычно созревает несколько фолликулов, а остальные растворяются. Доминантным является самый большой и развитый защитный элемент. Он защищает готовую к оплодотворению яйцеклетку. Непосредственно в период овуляции в правом или левом яичнике он способен достигнуть размера в несколько сантиметров. Под воздействием гормонов происходит его разрыв, в результате чего яйцеклетка высвобождается и устремляется к маточным трубам, а значит, появляется возможность беременности.
В редких случаях происходит одновременное созревание доминантов в обоих яичниках. Это дает возможность зачать двойню.
Существует также резерв фолликулов, которые готовы к оплодотворению. Их называют антральными. При подготовке к экстракорпоральному оплодотворению специалисты определяют, сколько фолликулов образуется, и на основании этих данных ставят прогнозы о вероятности беременности.
Стадии развития
За весь период жизни женщины, начиная с самого рождения, фолликулы проходят несколько стадий развития:
- Примордиальная стадия. Это незрелые фолликулярные клетки, которые закладываются во время формирования плода женского пола. Они очень малы и в диаметре не превышают 0,05 миллиметра. Фолликулы, способные к размножению делением, покрываются эпителием и переходят в следующую стадию.
- Первичные или преантральные образования достигают 0,2 мм в диаметре. Во время активного полового созревания девочки гипофизом активно синтезируется фолликулотропин, который ускоряет развитие клеток, укрепляет их мембраны и формирует защитный слой.
- Вторичные или антральные фолликулы в размерах увеличиваются до 0,5 мм. Их общее количество – около 8-10. Под воздействием эстрогенов внутренняя полость начинает заполняться жидкостью, которая растягивает стенки и провоцирует стремительный рост пузырьков. Вторичные фолликулы, кстати, считаются временными органами эндокринной системы, продуцирующими гормоны.
- В следующую стадию переходит, как правило, лишь одно фолликулярное образование – доминантное. Оно становится самым объёмным и заключает в себе практически полностью созревшую и готовую к оплодотворению яйцеклетку. Пузырёк состоит из большого количества гранулёзных клеток и призван обеспечивать надёжную защиту ооцита до момента овуляции. Остальные вторичные фолликулы в это время синтезируют эстрогены, обеспечивающие быстрое развитие главного пузырька.
- Третичный или предовуляторный пузырёк называется граафовым. Фолликулярная жидкость полностью заполняет его полость, её объём возрастает в сто раз по сравнению с исходным. Во время овуляции пузырёк разрывается, из него выходит яйцеклетка.
Нарушение развития фолликулов
Любые нарушения в развитии фолликулов приводят к серьезным последствиям, вплоть до бесплодия. Этому могут способствовать следующие отклонения:
- фолликулярные яичники;
- воспалительные процессы органов малого таза;
- недостаточная выработка женского гормона – эстрогена;
- нарушения эндокринной системы;
- проблемы с овуляцией;
- нарушения функций гипофиза;
- преждевременный климакс (хирургический или естественный);
- стрессы, депрессии, нервные перенапряжения.
Также очень важным моментом является состояние доминантного фолликула, который может отсутствовать, не достигнуть требуемых размеров, запоздать с созреванием или же не развиться совсем.
Нарушения фолликулярного аппарата
Одна из частых патологий — это созревание большого количества фолликулов, такое состояние называют — поликистоз яичников. При данной патологии не созревает доминантный фолликул, поэтому таким женщинам забеременеть очень трудно. Данная патология требует длительного гормонального лечения. Бывает, когда вырабатывается мало фолликулов, или они вообще не созревают. Это происходит из-за гормонального сбоя, когда в организме низкий уровень эстрогенов.
Персистирующие фолликулы
При возрастных изменениях или в подростковом возрасте нередко происходит нарушение деятельности фолликулярного аппарата, называемое персистенция. Главными симптомами заболевания являются нарушение менструации, обильные менструальные выделения, кровотечения. В этом случае происходит обратное развитие фолликула в яичниках, что может привести к образованию кисты. Чтобы киста не лопнула, назначается гормональная терапия. При значительных размерах опухоли лечение гормонами неэффективно, и требуется оперативное вмешательство. Явление персистенции сопровождается:
- гормональными нарушениями;
- утолщением слизистой эндометрия;
- отторжением эндометрия;
- сдавливанием матки;
- болевыми ощущениями внизу живота, сопровождающимися кровянистыми выделениями или кровотечением.
Плохо растут фолликулы
Недоразвитость структурных компонентов яичников является одной из основных причин женского бесплодия. При отсутствии доминантного фолликула в кровь не поступают лютеинизирующие гормоны, стимулирующее наступление овуляции. К основным причиной недостаточного развития половых клеток относятся:
- патологии гипоталамуса;
- нарушение генеративной функции яичников;
- злоупотребление противозачаточными средствами;
- последствия инфекции в малых половых органах;
- патологии щитовидной железы;
- эндокринные нарушения;
- депрессия и эмоциональная нестабильность;
- образование опухолей в гипофизе.
Адекватная гормональная терапия позволяет восстановить менструальный цикл и процесс созревания ооцитов. При неэффективности консервативного лечения назначается каутеризация яичников, которая заключается в хирургическом удалении недоразвитых клеток из половых желез.
Примордиальный фолликулез
Овариальный резерв (запас яйцеклеток у женщин) закладывается еще в утробе матери. Первичной стадией развития защитного фолликула является примордиальная. При этом зачатки яйцеклеток располагаются на внутренней поверхности яичников и защищены гранулезными клетками. Такая картина наблюдается вплоть до наступления менструации. Период полового созревания характеризуется:
- выработкой гормона, стимулирующего развитие фолликула:
- ростом ядра яйцеклетки под действием фолликулостимулирующего гормона;
- созреванием защитной оболочки яйцеклетки;
- ежемесячным развитием нескольких фолликулов, защищающих половую клетку.
Фолликулометрия
УЗИ-диагностику проводят с целью определения количества фолликулов, стадии их развития и роста.
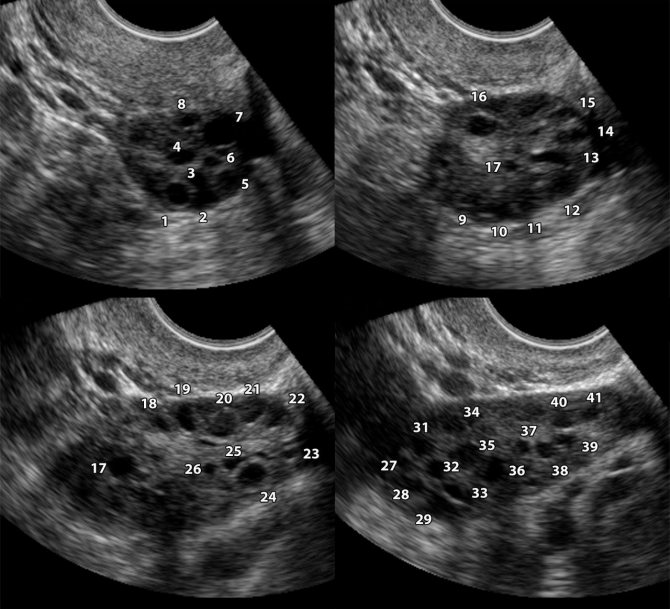
Поликистоз на снимках с фолликулометрии
Но проводить данное исследование в любой день нельзя, для правильной диагностики это делают в определённый день менструального цикла. Потому как в менструальную неделю определить их наличие просто невозможно. А вот на 8-9 день уже можно отчётливо увидеть наличие мелких «пузырьков».
Такое исследование позволяет определить, есть ли доминантные фолликулы, при этом в норме за 1 цикл развивается только один в одном из яичников. Но бывают случаи, когда их созревает несколько. Тогда повышается вероятность зачатия ребёнка и возникновения многоплодной беременности.
Доминантный фолликул на УЗИ находят по округлой форме и увеличенному размеру, нормальный размер в зрелом состоянии — до 20-25 мм.
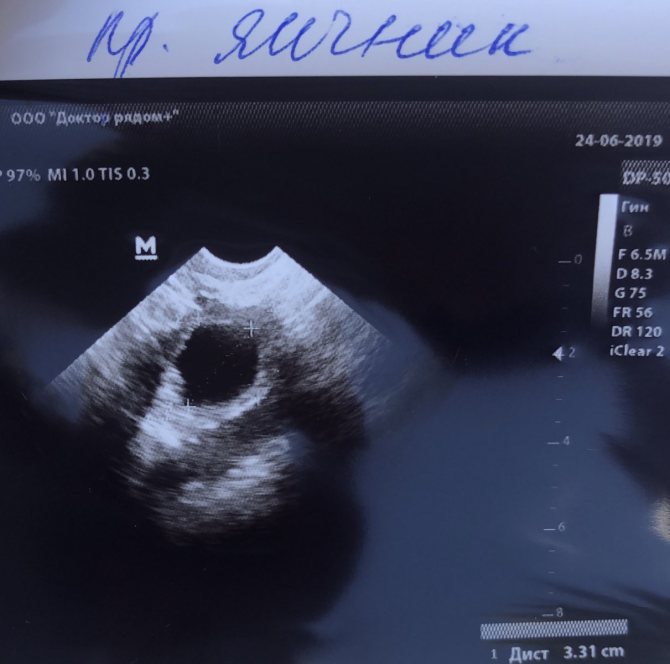
Доминантный фолликул на фолликулометрии
Антральный фолликулез
Антральные фолликулы в яичниках имеют размер не более 8 мм. Они развиваются на седьмой – восьмой день менструального цикла. Контроль их количества у женщин важен на стадии определения ее возможности забеременеть искусственным путем. Количество защитных элементов можно определить на УЗИ и по полученным данным определить резерв яйцеклеток, способных к оплодотворению.
Если антральные элементы достигают размера до 5 мм, вероятность наступления беременности низкая. При величине фолликулов 5 – 8 мм женщина с большой вероятностью забеременеет без помощи врачей. Стоит обратить внимание, что во время беременности фолликулы в яичниках не образуются.
Цели и преимущества метода
Ультразвуковое исследования фолликулогенеза показано женщинам с целью выявить возможные патологии процесса созревания яйцеклетки, а также для контроля качества биоматериала перед его забором для ВРТ. Исследование позволяет наблюдать рост доминантного фолликула и определить его готовность к оплодотворению.
Диагностика позволяет определить:
- Качество работы яичников и состояние репродуктивной системы женщины в целом;
- Состояние эндометрия;
- Выявить возможную причину бесплодия;
- Точный день цикла;
- Факторы, влияющие на сбой менструального цикла;
- Оптимальный день для проведения пункции фолликулов – актуально для женщин, участвующих в программах ЭКО или для доноров ооцитов.
Контроль фолликулогенеза, как метод диагностики, отличается следующими преимуществами:
- Является абсолютно безвредным и безболезненным (как обычное УЗИ малого таза).
- Может проводиться в трансвагинальном или трансабдоминальном формате.
- Отличается высокой точностью полученных результатов.
- Позволяет повысить шансы на успешное ЭКО или корректно определить причину патологий в репродуктивной системе женского организма.
- Имеет объективно доступную стоимость.
Преовуляторный фолликул
На последнем этапе своего созревания яйцеклетка готова к оплодотворению. При этом фолликул практически полностью наполнен жидкостью, за день до наступления овуляции возрастает выработка эстрогена и наблюдаются следующие явления:
- стимулируется выброс лютеина, способствующего наступлению овуляции;
- преовуляторный фолликул образует на своей стенке выпячивание, в месте которого позже происходит прорыв (овуляция);
- после наступления овуляции активизируется выработка прогестерона, предотвращающая отторжение эндометрия;
- образуется желтое тело, впоследствии формирующее сетку сосудов и способствующее появлению и развитию плаценты.
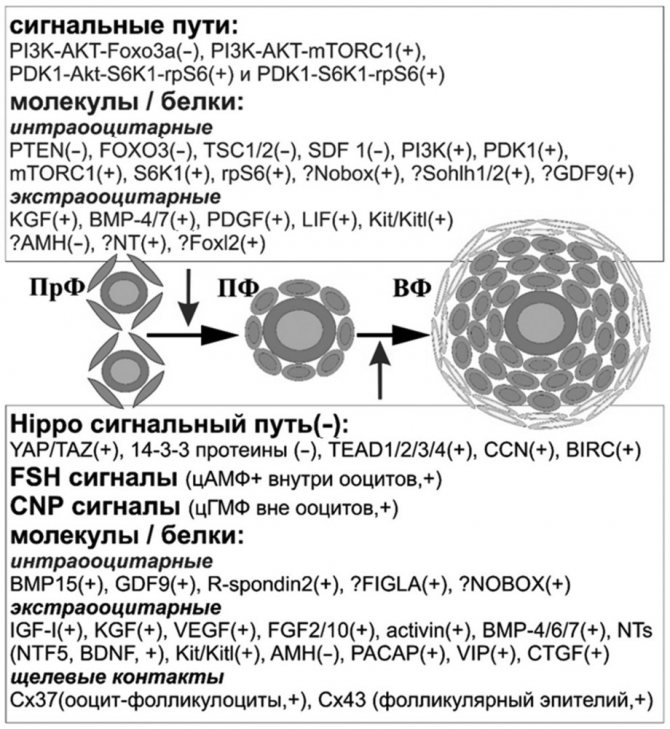
Схема основных факторов и сигнальных путей, вовлеченных в процесс раннего фолликулогенеза.ПрФ — примордиальные фолликулы, ПФ — первичные фолликулы, ВФ — вторичные фолликулы, (–) — поддержание «покоя» ПрФ/ограничение развития ПФ и ВФ, + — активация ПрФ/стимуляция роста ПФ и ВФ,? — потенциальное участие (составлено автором).
Продолжая обсуждение процессов фолликулогенеза, следует обратить внимание на основные и потенциальные факторы и сигнальные пути, вовлеченные в процесс раннего фолликулярного развития (см. схему).
Инициация фолликулярного роста
. Примордиальные фолликулы являются фундаментальными репродуктивными единицами яичника. Переход от состояния покоя до стадии роста называется рекрутированием, или активацией. Гистологически примордиальный фолликул содержит небольшой первичный ооцит (~ 25 мкм в диаметре), находящийся на стадии профазы I мейоза, окруженный одним слоем уплощенных фолликулярных клеток, тесно соединенных с ооцитом и базальной мембраной. Примордиальные фолликулы находятся в микросреде, исключающей контакт с большинством других клеток и кровеносными сосудами, имеют ограниченный доступ к эндокринной системе. Гистологическими признаками активации покоящихся фолликулов служит приобретение митотического потенциала клетками фолликулярного эпителия (изменение формы на кубическую). Процесс сопровождается ростом ооцита [1].
Рекрутирование примордиальных фолликулов в основном контролируется паракринной взаимосвязью между ооцитом, фолликулярным эпителием, соседними текальными и интерстициальными клетками. Пул покоящихся фолликулов находится под строгим контролем ингибирующих факторов, активация происходит при увеличении стимулирующего влияния. Последовательность действий во многом опосредована секретируемыми ростовыми факторами, включая несколько членов суперсемейства трансформирующего фактора роста-бета (TGF-β) [2, 3].
Экспериментальные данные позволяют предположить, что не только внешние стимулы, но и сами ооциты в значительной степени управляют передачей сигналов, регулирующих набор и скорость роста фолликулов. Основные сигнальные пути проходят через PI3K (фосфатидилинозитол-3-киназу) [4].
Во-первых, для сдерживания активации фолликулов требуется сигнализация PI3K-AKT-Foxo3a [1, 3, 5]. Данный сигнальный путь опосредуется PTEN (фосфотензин) [6]. Отсутствие PTEN активирует PI3K с образованием фосфолипида фосфатидилинозитол-3, -4, -5-трифосфата (PIP3) и Akt (протеинкиназа B) сигнализацию. На PTEN-дефицитных мышах (OoPten –/–) показана сверхактивация примордиальных фолликулов (рост достигает стадии первичных и вторичных) и раннее истощение яичников [7]. Белок Foxo3a (for khead box O3A), принадлежащий к for khead семейству транскрипционных факторов, также сдерживает рекрутирование. Foxo3a обнаруживается в ядрах ооцитов и фолликулярных эпителиоцитов примордиальных фолликулов. В растущих фолликулах экспрессия фактора снижается. Foxo3a-нокаутированные мыши (Foxo3a –/–) демонстрируют схожий с OoPten –/– фенотип, сопровождающийся ростом ооцитов и активацией фолликулов [5]. Истощение пула примордиальных фолликулов у Foxo3a –/– приводит к бесплодию. У OoPten –/– животных постоянная Akt активация подавляет Foxo3a, поэтому PTEN в указанном сигнальном пути находится выше FOXO3 [7].
Вторым сигнальным путем является PI3K-AKT-mTORС1 (протеинкиназа-серин-треониновой специфичности) [8]. Мультимолекулярный сигнальный комплекс TORC1 (мишень рапамицина млекопитающих) контролируется гетеродимерными белками: TSC1 (гамартин) и TSC2 (туберин) [1, 3]. У TSC1- и TSC2-дефицитных мышей (OoTsc1 –/–, OoTsc2 –/–) отмечается преждевременная активация примордиальных фолликулов за счет увеличения сигнализации S6K1-rpS6 (S6 киназа 1 и рибосомальный протеин S6). Фенотип также аналогичен OoPten –/– [9]. Оба белка (TSC и PTEN) регулируют рекрутирование фолликулов по пути S6K1-rpS6. TSC ингибирует фосфорилирование S6K1 до треонина 389, PTEN — до треонина 229. Таким образом, внутриклеточная (интраооцитарная) сигнализация, поддерживающая «покой» примордиальных фолликулов, обеспечивается тремя основными факторами: PTEN, FOXO3 и TSC.
Активация PI3K также обусловлена внеклеточными сигналами. Паракринная сигнализация между ооцитом и соматическими клетками окружения обеспечивается основным лиганд-рецептором — RPTK (рецептор тирозинкиназы). В постнатальных яичниках Kit (стволовой клеточный фактор) экспрессируется на поверхности ооцитов, а Kitl (лиганд) обнаруживается в прегранулезных клетках и фолликулярных эпителиоцитах всех типов растущих фолликулов [10].
На ранних стадиях развития фолликулов, в которых FSH-рецепторы отсутствуют, передача сигналов PI3K контролируется сигнализацией Kit/Kitl. В первичных ооцитах Kitl приводит к фосфорилированию Akt и Foxo3a. У млекопитающих Akt инактивирует Foxo3a через внеклеточную сигнализацию с участием факторов роста и инсулина. Эффект последнего для активации примордиальных фолликулов заключается в ингибировании FOXO3. В сигнальном пути PI3K PIP2 (фосфатидилинозитол-4,5-бисфосфат) фосфорилируется до PIP3. Фактор является вторичным мессенджером для PDK1 (3-фосфоинозитидзависимой киназы-1) и Akt. Последняя способствует жизнеобеспечению клеток, вызывает рост и активацию ооцитов. У мышей с ооцитспецифическим исключением гена, кодирующего PDK1 (OoPDK1 –/–), подавление сигналов PDK1-Akt-S6K1-rpS6 приводит к истощению пула примордиальных фолликулов [7]. В противоположность этому PTEN ингибирует PI3K путем превращения PIP3 в PIP2. Главной мишенью Akt является семейство FOXO, подавляющее активацию примордиальных фолликулов и поддерживающее фолликулы в состоянии покоя, стимулирующее проапоптотические гены. Однако при двойном нокауте (OoPten –/–, OoPDK1 –/–) деактивация фосфорилирования rpS6 предотвращает чрезмерную фолликулярную активацию в отличие от такового при изолированном удалении PDK1 (OoPDK1 –/–) [7].
В дополнение к двум сигнальным путям ауто-, паракринным действием, ингибирующим активацию фолликулов, обладает хемокин подсемейства CXC, вероятно, выделяемый ооцитами — SDF1 (стромальный клеточный фактор 1). CXCL12 и его рецептор — CXCR4 участвуют в различных физиологических процессах, в том числе и в миграции примордиальных герминогенных клеток гонад. Экспрессия SDF1α увеличивается при развитии фолликулов, а культивация неонатальных яичников in vitro
с ингибитором фактора (AMD3100) сохраняет фолликулы в неактивном состоянии, поэтому сигнализация SDF1/CXCR4 может играть важную роль в поддержании пула примордиальных фолликулов [11].
В рекрутировании примордиальных фолликулов также участвуют факторы, находящиеся вне непосредственного взаимодействия комплекса ооцит—фолликулоциты. Стромально-интерстициальные клетки, окружающие примордиальный фолликул, секретируют KGF (фактор роста кератиноцитов), BMP4 и BMP7 (костные морфогенетические белки 4 и 7), PDGF (тромбоцитарный фактор роста), LIF (ингибирующий лейкоз фактор). Факторы способствуют переходу примордиальных фолликулов в растущий пул [1, 10]. BMP4 и BMP7 являются членами суперсемейства TGFβ, связываются с двумя типами рецепторов серин-треонин киназы (BMPRII/ActRIIA и ALK3/ALK6). Сигнальная трансдукция проходит через SMAD 1/5/8 путь [12]. Белки активно экспрессируются в клетках теки антральных фолликулов, в то время как рецепторы обнаруживаются в фолликулярном эпителии и ооцитах. Наряду с фолликулярной активацией известными функциями BMP4 и BMP7 являются содействие овуляции, синтез ароматазы клетками гранулезы, содействие секреции эстрогенов и ингибирование прогестерона в антральных фолликулах [13].
KGF, PDGF и LIF повышают регуляцию Kitl в клетках гранулезы. В примордиальных фолликулах FGF7 и PDGF экспрессируются в ооцитах и стромальных клетках окружения (паттерны экспрессии у человека и грызунов различаются [14]), а LIF — в прегранулезе и соматических клетках [10]. Несмотря на участие представленных факторов в активации примордиальных фолликулов их критическая роль в обеспечении репродуктивной функции должна быть уточнена. Например, у KGF-дефицитных мышей (FGF7 –/–) дефекты рождаемости не выявлены. Нокаут LIF (LIF –/–) вызывает нарушение имплантации, но в целом отмечаются нормальный фолликулогенез и овуляция [10]. Способность многих молекул влиять на Kitl экспрессию, вероятно, является следствием эволюционно сложившихся «избыточных» путей передачи сигнала и обеспечивает компенсацию одних факторов при дефиците других.
Факторами транскрипции множественных сигнальных путей, регулирующими первый шаг фолликулогенеза, являются также Foxl2, GDF9, Nobox, SOHLH1, АМГ и нейротрофины.
Помимо FOXO3, другим фактором fork head семейства, играющим ключевую роль в переходе примордиальных фолликулов в растущий пул, является Foxl2 (For khead box L2). У мышей белок экспрессируется в прегранулезных клетках, выражение Foxl2 снижается в преантральных фолликулах. Фактор имеет решающее значение при переходе фолликулярного эпителия от плоскоклеточного до кубического при начале роста фолликулов. У Foxl2-дефицитных животных (Foxl2 –/–) примордиальные фолликулы образуются, однако они не развиваются до вторичных в результате дефекта дифференцировки клеток гранулезы. Большая часть ооцитов активируется преждевременно, о чем свидетельствует экспрессия GDF9 (фактор роста/дифференцировки 9). Блок пролиферации замедляет рост ооцитов и приводит к атрезии фолликулов [15].
GDF9 является одним из ведущих факторов роста ооцитарного происхождения, обеспечивающим функции соматических клеток окружения [16]. Белок непосредственно регулируется Nobox. GDF9 экспрессируется в ооцитах растущих фолликулов. Лечение неполовозрелых крыс с рекомбинантным GDF9 приводит к увеличению скорости фолликулярной активации и роста фолликулов. Нокаут фактора у мышей (GDF9 –/–) вызывает рост ооцитов, обусловленный повышенной регуляцией Kitl (увеличение сигнализации Kitl/Kit, повышение экспрессии Kit). В то же время резко возрастает уровень ингибина α, приводящий к снижению пролиферации фолликулоцитов, отсутствию условий для образования текальной оболочки. В результате GDF9 –/– фолликулы далее первичной стадии не формируются. Результаты свидетельствуют о том, что GDF9 играет роль в наборе фолликулярных клеток и в то же время ограничивает рост ооцитов [10]. Паттерны экспрессии, которые выявляются в основном в ооцитах растущих фолликулов, демонстрируют роль фактора скорее в процессах дальнейшего развития фолликулов, чем в рекрутировании примордиальных фолликулов.
Nobox (новорожденный ооцитспецифический гомеобоксный протеин), Sohlh1 и Sohlh2 (сперматогенез- и оогенезспецифические белки 1 и 2, построеннные по принципу «спираль—петля—спираль») являются критическими факторами транскрипции перехода фолликулов из примордиального в растущий пул. Все три белка обнаруживаются в кластерах герминогенных клеток, примордиальных и первичных фолликулах. Sohlh1 и Sohlh2 быстро исчезают, как только ооциты достигают вторичной стадии развития, Nobox определяется во всех стадиях фолликулогенеза. Мыши с недостатком какого-либо фактора транскрипции являются стерильными. Несмотря на то что яичники от новорожденных нокаут-особей Nobox –/– или Sohlh1 –/– содержат сходное с контролем количество клеточных кластеров герминногенных клеток и примордиальных фолликулов, рост фолликулов далее примордиальной стадии нарушается. Sohlh2-дефицитные животные (Sohlh2 –/–) имеют похожий Nobox –/– или Sohlh1 –/– фенотип, но иногда формируют фолликулы с несколькими слоями гранулезы [3, 10]. В отличие от молекулярно-биологических изменений в Nobox –/– яичниках, у новорожденных мышей Sohlh1 –/– или Sohlh2 –/– отмечается даун-регуляция Kit рецептора, что подчеркивает избыточность сигнальных путей для формирования примордиальных фолликулов в пренатальном периоде и их активации в постнатальном.
АМГ (антимюллеров гормон) и его рецептор AMнRII у человека и грызунов выражаются в фолликулоцитах первичных и преантральных фолликулов. Роль АМГ в изначальном фолликуле была исследована в Amh –/–. Несмотря на то что самки мышей, лишенные AMГ, плодородны, отмечается преждевременное истощение примордиальных фолликулов (АМГ ингибирует их рост). При культивировании неонатальных яичников мыши (равно как и ткани яичников человека) в присутствии рекомбинантного АМГ количество растущих фолликулов снижается в среднем в 2 раза. В настоящее время АМГ является единственным экзогенным негативным регуляторным фактором фолликулярной активации [17]. У человека установить прямую связь между концентрацией АМГ и пулом примордиальных фолликулов трудно (у мышей существует сильная корреляция между указанными параметрами). Тем не менее у женщин АМГ положительно коррелирует с количеством антральных фолликулов [3, 10].
NT (нейротрофины) являются растворимыми факторами роста, функции которых выходят за пределы нервной системы и включают регулирование раннего фолликулогенеза. По крайней мере, 4 из 5 NT, а также их рецепторы выражены в яичнике: NGF (фактор роста нервов), BDNF (нейротропный фактор мозга), NTF3 и NTF5 (нейротрофины 3 и 5). NT экспрессируются в соматических клетках и ооцитах фолликулов до начала их роста. В яичниках NGF-дефицитных животных (NGF –/–) обнаруживаются в основном примордиальные фолликулы, популяция первичных фолликулов немногочисленна. Фенотип гонад аналогичен Foxl2 –/–, а передача сигналов NGF также имеет важное значение в дифференциации фолликулоцитов от плоскоклеточной до кубической формы при активации фолликулов. Несмотря на то что мыши, лишенные общего р75 рецептора NGF, способны к размножению без дефектов фолликулярной формации, дефицит NTRK1 (рецептор тирозинкиназы высокого сродства NGF) вызывает перинатальную летальность, а овариальные эффекты NTRK1 остаются неизвестными [3, 10].
Базальный фолликулярный рост (рост фолликулов от инициации до стадии малых антральных — 2 мм в диаметре). На ранних этапах развития первичный фолликул представлен кубическими фолликулоцитами, формирующими один слой вокруг ооцита (диаметр которого ~ 25 мкм). На протяжении преантрального фолликулогенеза структура фолликула изменяется, главным образом, за счет пролиферации фолликулярных клеток, формирующих уже несколько слоев, а также приобретения теки. Форма фолликулоцитов становится цилиндрической, размеры ооцитов продолжают увеличиваться. Основные события, которые происходят в первичном фолликуле, включают экспрессию рецептора ФСГ (фолликулостимулирующий гормон), рост и дифференцировку ооцитов. Рецепторы ФСГ в фолликулярном эпителии первичных фолликулов обнаруживаются вскоре после инициации фолликулярного роста. Ключевыми стимуляторами экспрессии рецептора ФСГ являются собственно ФСГ, активин, цАМФ (циклический аденозинмонофосфат) и TGF. Несмотря на то что рекрутирование и начальные стадии роста фолликулов не зависят от гонадотропинов, ФСГ необходим для развития первичного фолликула до преантральной стадии. A. Hsueh и соавт. [18] предлагают использовать два термина в обозначении преантральной и антральной стадий роста фолликулов — «ФСГ-чувствительный» и «ФСГ-зависимый» периоды. В результате реактивации генома ооцита в период от примордиальной до преантральной стадии размер половой клетки увеличивается в 5 раз, окружающий внеклеточный матрикс образует блестящую оболочку (zona pellucida
) [1].
Центральными регуляторами белков zona pellucida
(zp1, zp2, zp3) являются транскрипционные молекулы FIGLA (фактор герминогенной линии α), Nobox и факторы роста GDF9, BMP15 [1].
Zp-дефицитные мыши являются бесплодными или субфертильными, так как отсутствуют рост и дальнейшее развитие фолликулов. FIGLA же необходим для формирования примордиальных фолликулов пренатально [19]. При удалении фактора транскрипции Figα (Figla –/–) количество кластеров герминогенных клеток аналогично контролю, однако в постнатальном периоде примордиальные фолликулы отсутствуют. Figla –/– яичники демонстрируют снижение активности 165 генов и увеличение активности 38 генов, большая часть которых кодирует транскрипционные факторы обязательных функций нуклеиновых кислот [10].
Наибольшая роль в регуляции преантрального фолликулогенеза отводится ооцитсекретируемым факторам, в первую очередь GDF9 и BMP15 [20, 21]. Оба фактора вырабатываются ооцитами, стимулируют пролиферацию клеток гранулезы и развитие теки [10, 22]. Нарушение регуляции GDF9 отмечено при синдроме поликистозных яичников, мутация BMP15 сопряжена с преждевременной недостаточностью яичников [1]. Значение GDF9 и его регулятора Nobox в базальном росте фолликулов представлено выше.
BMP15 относится к суперсемейству TGFβ [23]. Аминокислотный состав фактора более чем наполовину идентичен GDF9, в силу чего паттерны экспрессии обоих белков аналогичны. Однако BMP15 не требуется во время преантрального фолликулогенеза у мышей. BMP15 –/– животные субфертильны, так как уменьшается количество фолликулов более поздних стадий развития и желтых тел за счет потери ооцитов. В то же время соотношение значимости GDF9 и BMP15 в процессах раннего и позднего фолликулогенеза у различных видов млекопитающих может различаться. Например, у BMP15 –/– овец показан аналогичный мышиному GDF9 –/– фенотип яичников, что демонстрирует критическую роль BMP15 именно в процессах раннего фолликулогенеза, избыточность регуляторных путей и, вероятно, эволюционные особенности. Экспрессия BMP15 регулируется ФСГ по средствам сигнализации Kit. Помимо мощной митогенной функции, для фолликулоцитов GDF9 и BMP15 также ингибируют апоптоз и преждевременную лютеинизацию, стимулируют энергетический и холестериновый метаболизм, участвуют в экспансии клеток кумюлюса. На более поздних этапах фолликулогенеза в фолликулярных эпителиоцитах факторы взаимодействуют с рецепторами морфогенетического белка II и киназоподобным рецептором активина, стимулируют SMA- и MAD-связанную внутриклеточную сигнализацию, участвуя в созревании ооцита [24].
Третьим ооцитарным фактором пролиферации фолликулярного эпителия является R-spondin2. Белок является фактором роста стволовых клеток, действует через Wnt сигнальный путь. Участие R-spondin2 в преантральном фолликулогенезе впервые продемонстрировано Y. Cheng и соавт. [25]. В естественных условиях введения агониста R-spondin стимулирует развитие фолликулов до преантральной стадии как у неполовозрелых мышей, так и у десенситизированных введением антагониста ГнРГ взрослых особей.
В дополнение к ооцитарным факторам, большая группа белковых лигандов, секретируемых клетками гранулезы и действующих через RTK (рецепторные тирозинкиназы), модулирует рост фолликулов: IGF1, KGF, VEGF, FGF2, FGF10, активины, BMP6, АМH, PACAP, VIP, CNP, CTGF. Овариальная роль РТК-опосредованной сигнализации некоторых лигандов представлена выше, ключевые другие обсуждаются ниже. Однако физиологическое значение указанных факторов в преантральном фолликулогенезе необходимо, но менее очевидно [18].
После активации примордиальных фолликулов Kit/Kitl сигнализация сохраняется и является также критически важной для роста ооцитов и организации текаклеток вокруг фолликулов [26]. Вероятно, несколько факторов влияют на рост фолликулов, стимулируя фолликулоциты увеличивать экспрессию Kitl [27]. Повышенная экспрессия Kitl в фолликулярном эпителии усиливает воздействие на ооциты, демонстрируя в дальнейшем «прямое» паракриновое взаимодействие между половой и соматическими клетками окружения [1, 10].
Стимулирующие функции нейротрофинов также поддерживаются во время преантрального фолликулогенеза. Сигналы NTF5 и BDNF на ооцит через NTRK2 при росте первичных фолликулов, вероятно, имеют избыточный характер. Потеря Ntf5 не изменяет количество преантральных фолликулов, однако совместная инактивация Ntf5 и BDNF значительно снижает их число, формируя фенотип, аналогичный Ntrk2 –/–. Действие NTF5 и BDNF не зависимо от сигнальных путей, связанных с Gdf9 и Kitl [10].
Важным событием в развитии первичного фолликула является образование интимных межклеточных связей между ооцитами и фолликулоцитами. Оба типа клеток формируют многочисленные цитоплазматические выступы и микроворсинки, распространяющиеся друг на друга для увеличения площади соприкосновения. Некоторые микроворсинки эпителиоцитов достигают ядерной мембраны половой клетки. Многочисленные щелевые контакты (межклеточные каналы) представлены белками — коннексинами (Cx), обеспечивают диффузию ионов, метаболитов и сигнальных молекул. В яичниках млекопитающих экспрессируются несколько Cx, по крайней мере два играют существенные и различные роли в процессе фолликулогенеза. Cx37 (GJA4) является преобладающим щелевым белком, синтезируемым ооцитами после фолликулярной активации. Фолликулоциты, наоборот, в основном производят Сх43 (GJA1). Между ооцитами и клетками гранулезы образуются гетеротипические щелевые соединения, состоящие из обоих Cx, в то время как между фолликулярными эпителиоцитами — гомотипические, представленные только Cx43. Значимость взаимодействия через щелевые контакты была продемонстрирована на грызунах. Сх43-нокаутированные мыши (Gja1 –/–) погибают в раннем постнатальном периоде из-за сердечных пороков развития. Яичники новорожденных особей маленькие, что обусловлено блоком начальной стадии развития фолликулов, нарушением пролиферации фолликулоцитов и замедленным ростом ооцитов с дефектами мейотического созревания. Cx37-дефицитные яичники (GJA4 –/–) также содержат некомпетентные ооциты, рост фолликулов прогрессирует только до поздней преантральной стадии. В гонадах обнаруживаются структуры, напоминающие желтые тела. Соответственно, связь через Cx37 является основным механизмом регулирования образования желтых тел, а при нарушении взаимодействия ооцит—фолликулоциты происходит преждевременная лютеинизация фолликулов [1, 10].
После приобретения фолликулом двух слоев фолликулоцитов развивается морфологически различимый слой соматических клеток. Мезенхимальные клетки, обнаруживающиеся вокруг базальной мембраны, вырабатывают BMP4 [28]. Экспрессия фактора в стромальных клетках, вероятно, необходима не только для активации примордиальных фолликулов, функциональный маркер является условием для дифференцировки клеток теки. Окружающие фолликулоциты интерстициальные клетки (внутренняя тека) имеют ультраструктурные особенности: большое количество митохондрий с трубчатыми кристами, гладкую эндоплазматическую сеть, а также многочисленные липидные везикулы, соответствующие основной их функции, являются источником андрогенов для синтеза эстрогенов в клетках гранулезы. Наружная тека представлена фибробластами, гладкомышечными клетками и макрофагами, имеет важное значение во время овуляции. Формирование теки во время преантрального фолликулогенеза является гонадотропиннезависимым, так как клетки–предшественники лишены рецепторов ЛГ, а слой теки образуется в яичниках ФСГ-дефицитных мышей. Дальнейшее развитие текальных оболочек сопровождается новообразованием многочисленных мелких кровеносных сосудов. Следовательно, питательные вещества и гонадотропины с током крови циркулируют вокруг развивающегося фолликула. Перед вступлением фолликула в антральную стадию роста последний содержит пять гистологически различных, но взаимодействующих структурных единиц: зрелый ооцит, окруженный zona
pellucida
, около 9 слоев фолликулярных эпителиоцитов и 2 слоя теки с капиллярной сетью кнаружи от базальной мембраны [1, 10].
Факторы, регулирующие текальную дифференцировку, окончательно не установлены, предполагается, что это небольшие молекулы 20—25 кД, секретируемые растущими фолликулами. Потенциально способствующие дифференцировке текаклеток маркеры включают ИФР (инсулиноподобные факторы роста), Kitl и GDF9. При культивировании недифференцированных тека-интерстициальных клеток крыс в искусственной среде с мРНК для рецепторов ЛГ/хориогонадотропин (Lhcgr) и Cyp17a1 начиналась продукция андрогенов. IGF1 повышает экспрессию Lhcgr, Cyp11a1 и 3β-гидроксистероиддегидрогеназы (Hsd3b1), а при добавлении Kitl выявляются мРНК других маркеров — Star (стероидогенного острого регуляторного белка) и Cyp17a1 [1, 10].
Некоторые эффекты действия GDF9 на развитие теки являются косвенными [29]. Gdf9 –/– яичники демонстрируют отсутствие маркеров Cyp17a1, Lhcgr и Kit, несмотря на большое количество предшественников текальных клеток в интерстиции и высокие концентрации ФСГ и Л.Г. Рекомбинантный GDF9 у мышей регулирует IGF1 в культивируемых клетках гранулезы. Действие фактора может быть еще более важным в дифференцировке текаклеток, так как двойной нокаут ингибина α и GDF9 сопровождается формированием морфологически различимого слоя внутренней теки, однако лишенного большинства указанных выше маркеров их активности.
В антральной гонадотропинзависимой стадии роста ЛГ-стимуляция приводит к экспрессии ключевых ферментов стероидогенеза в текаклетках: CYP11A1, HSD3B1 и CYP17A1. ЛГ также повышает регуляцию Star, обеспечивающего доставку холестерина к внутренней митохондриальной мембране, где находится CYP11A1. Фолликулоциты реагируют на ФСГ повышенной регуляцией CYP19A1 и 17b-гидро-ксистероиддегидрогеназы (HSD17B1). Так как эстрадиол имеет существенное значение для более поздних стадий фолликулогенеза и овуляции, а текальные эндокриноциты экспрессируют рецептор ЛГ задолго до формирования фолликулярной полости, необходимо подавление избыточного биосинтеза андрогенов в преантральных и малых антральных фолликулах. Фолликулярный эпителий секретирует активины, модулирующие эффекты действия ЛГ на клетки теки. Kitl фолликулоцитов также регулирует факторы теки, оказывающие аутокринное тормозящее действие на производство андростендиона: TGFβ, TGFα, FGF7 и HGF (фактор роста гепатоцитов) [1, 10].
Окончание преантрального фолликулогенеза сопровождается такими изменениями ооцитов, при которых возможно самопроизвольное возобновление мейоза при его удалении из фолликулярной среды. Однако известно, что во время фолликулогенеза в ооцитах редко активируется мейоз. Решающую роль в регулировании мейотической компетенции ооцитов играет цАМФ, подавляющий возобновление мейоза [1]. Действие ФСГ опосредуется преимущественно сигнализацией цАМФ внутри самого ооцита. Помимо ФСГ, CNP (натрийуретический пептид С-типа) является паракринным фолликулостимулирующим фактором, в том числе на преантральной стадии. CNP активирует NPR2 (рецептор, гуанилатциклаза В), обеспечивающий сигнализацию вторичного мессенджера – цГМФ (циклический гуанозинмонофосфат). Существует доказательство того, что именно цГМФ (а не цАМФ) переходит через щелевые контакты из фолликулоцитов в ооцит, где ингибирует функцию PDE3A (фосфодиэстеразы 3A), предотвращая PDE3A опосредованное расщепление цАМФ и сдерживая, таким образом, мейоз [18].
В течение почти 100 лет было известно, что повреждение яичников может способствовать фолликулярному росту [18], однако только в настоящее время появилось убедительное научное обоснование указанного положения. Пионерами создания нового понимания раннего фолликулогенеза являются ученые Стенфордского университета во главе с A. Hsueh. Исследователи показали, что сигнальный путь Hippo ограничивает, а CNN факторы роста стимулируют преантральный фолликулярный рост [18].
Hippo является ведущим эволюционным сигнальным путем, контролирующим размеры органа во всех многоклеточных организмах, ингибирует пролиферацию клеток и стимулирует апоптоз (определяет дальнейшее развитие стволовых/прогениторных клеток) [30]. Сигнализация состоит из нескольких негативных регуляторов роста, действующих на каскад серин/треонин киназы, в конечном итоге фосфорилирует и инактивирует ключевые транскрипционные коактиваторы YAP и TAZ (эффекторы Hippo-супрессорного пути, обладающие пролиферативной и онкогенной активностью). При активации сигналов Hippo коактиваторы секвестированы в цитоплазме различными белками (14−3-3 протеины), способствующими их деградации. При нарушении передачи сигналов Hippo ядерные YAP/TAZ взаимодействуют с фактором транскрипции TEAD фолликулярных эпителиоцитов, увеличивают экспрессию факторов роста (CСN) и ингибиторов апоптоза (BIRC). Сигнальный путь Hippo регулируется физическим и механическим микроокружением клеток. Механические сигналы от внеклеточного матрикса, участков клеточной адгезии, состояния актомиозинового цитоскелета сходятся вместе в пути Hippo и влияют на дальнейшую судьбу клеток [31]. Повреждение гонад (экстрацеллюлярного матрикса) полимеризует G-актин до F-актина, нарушает Hippo сигнализацию — активирует YAP/TAZ.
После активации и выхода из состояния покоя локальные сигналы Hippo определяют различную траекторию роста фолликулов, связанную с этим разную продолжительность их жизни (23—90 дней) [18]. Примордиальные фолликулы расположены в кортикальной области, экстрацеллюлярный матрикс которой более жесткий, чем таковой в медуллярной области. Культивирование мезенхимальных стволовых клеток в жестком внеклеточном матриксе увеличивает YAP активность, в то время как в мягкой матрице действие YAP снижается [32]. Одним из вероятных механизмов поддержания фолликулов в неактивном состоянии (наряду с внутриооцитарными сигнальными путями) является механотрансдукция Hippo, так как известно, что с увеличением роста фолликулов они отдаляются от поверхности яичников, где активность YAZ повышается. Более крупные фолликулы усиливают сигнализацию Hippo в соседних небольших фолликулах, подавляя их рост. Во время каждой овуляции выраженные структурные изменения, обусловленные разрывом фолликула, могут нарушать местную сигнализацию Hippo — индуцировать полимеризацию актина вблизи поверхности яичников. Ежемесячное нарушение сигнализации, приводящее в итоге к «сверхпролиферации» поверхностного целомического эпителия, может способствовать формированию рака яичников. Дефекты генов сигнального пути Hippo сопряжены не только с онкогенезом (LATS ½), механотрансдукция также имеет место при первичной овариальной недостаточности (DIAPH2, BIRC1), синдроме поликистозных яичников (LATS ½) и снижении овариального резерва при бесплодии (DIAPH2, DIAPH3, CCN2) [18].
Таким образом, регуляция раннего фолликулярного роста имеет сложный механизм, включающий несколько сигнальных путей, интра- и экстраооцитарные факторы. Несмотря на проведенные исследования, все еще не определены механизмы инициации базального фолликулярного роста, а также факторы, регулирующие текальную дифференцировку. Однако не остается сомнения в том, что представленные регуляторные механизмы являются основой сохранения овариального резерва. Подробное изучение сигнальных путей регуляции фолликулярного роста позволит получить новые возможности для профилактики и лечения дисфункции яичников, что является основной задачей репродуктивной биологии и медицины.
Авторы заявляют об отсутствии конфликта интересов.
Какие нарушения могут быть обнаружены?
УЗИ поможет обнаружить следующие патологические состояния:
- остановка роста фолликула, его обратное развитие и не наступление овуляции;
- фолликул не разрывается, соответственно, не наступает выход яйцеклетки;
- фолликулярная киста — состояние, при котором в фолликуле может накапливаться жидкость;
- полное отсутствие развития фолликула, что может быть вызвано нарушением гормонального фона.
| УЗИ женской половой сферы | Цена (руб.) |
| Фолликулогенез с ЦДК (1 обследование) | 350 |
| Трансвагинальное УЗИ матки и придатков с ЦДК | 700 |
| Трансабдоминальное УЗИ матки и придатков с ЦДК | 450 |
| Трансвагинальное УЗИ для определения беременности | 700 |
| Цервикометрия | 350 |
| УЗИ уретры | 550 |
Исследование фолликулогенез — важный шаг в подготовке к естественному зачатию и наступлению беременности. Чтобы записаться на диагностику в клинику «Алтайский медицинский центр», вы можете воспользоваться онлайн формой на сайте или позвонить по телефонам: (3852)222-250 и (3852)222-250.
Записаться на приём